عصارة خلوية
العُصارَة الخَلَوِيَّة[3][4][5][6][7][8] أو عُصارة الخَليَّة[3][5][9] (بالإنجليزية: Cytosol) وتُترجم حرفيًا إلى[10] «السيتوسول»[11]، وتُسمى أيضًا المَصفُوْفَة السيتوبلازمية أو المَطْرِس السيتوبلازمي[lower-alpha 1] (بالإنجليزية: Cytoplasmic matrix)، هيَ المادة الرئيسية للسيتوبلازم، وهي سائل مُعلق موجود داخل الخلايا، حيثُ تُعلق أو تذوبُ فيه المواد الكيميائية العضوية وغير العضوية والجزيئات والصبغيات والبلورات والعضيات الخَلوية.[14] تشكل العصارة الخلوية مُعظم السائل داخل الخلية، وتقسم إلى أكثر من حيز بواسطة الأغشية، كما هو الحال في المَصفُوفَة الميتوكندرية التي تُقسم الميتوكندريون إلى العديد من الحُجرات.
| العُصارَة الخَلَوِيَّة | |
|---|---|
| الاسم اللاتيني Cytosol; Matrix cytoplasmica | |
 مُخطط الخلية الحيوانية النموذجية (العُصارة الخلوية رقم 11) | |
| تفاصيل | |
| نوع من | كيان تشريحي خلوي [1] |
| جزء من | سيتوبلازم [1] |
| معرفات | |
| ترمينولوجيا هستولوجيكا | H1.00.01.0.00008 |
| FMA | 66836 |
| ن.ف.م.ط. | A11.284.430.214.200، وA11.284.430.429.200، وA11.284.835.450.200 |
| ن.ف.م.ط. | D003600 |

تتواجد العُصارة الخَلَويَّة في الخلايا حقيقيات النوى ضِمن الغشاء الخلوي وتُشكل جزءًا مِن السيتوبلازم الذي يضُم أيضًا الميتوكندريا والبلاستيدات والعُضيات الخلوية الأخرى (ولكن لا يضم هياكلها وسوائلها الداخلية)، كما أنَّ نواة الخلية تكون مُنفصلة، وبالتالي فإنَّ العُصارة الخَلَويَّة هي عبارة عن مَصفوفة سائِلة مُحيطة بالعضيات الخلوية، وفي حَقيقيات النوى تحدث العديد مِن المسارات الأيضية في العُصارة الخلوية، ويحدُث جزء آخر داخل العُضيات،[15] أما في بدائيات النوى، فإنَّ مُعظم التفاعلات الكيميائية لِعملية الأيض تحدث في العُصارة الخلوية، وجزء بسيط مِنها يحدث ضمن الأغشية أو في الحيز المُحيط بالجِبلَة المُحيطيَّة.[16]
العُصارَة الخَلَوِيَّة هي خليط معقد من المواد المذابة في الماء،[17] وعلى الرغم من أنَّ الماء يُشكل الغالبية العظمى من العُصارة إلا أنَّ خواصه وتركيبه ضمن الخلايا غير مفهوم جيدًا.[18] يختلف تركيز أيونات الصوديوم والبوتاسيوم في العُصارة الخلوية عن السائل خارج الخَلية، وهذا الاختلاف مُهم في بعض العمليات مِثل التنظيم الأسموزي وتأشير الخلية وتوليد جهد الفعل في الخلايا المستثارة مثل الخلايا العضلية والعصبية والصماء. تحتوي العصارة الخلوية أيضًا على كمياتٍ كبيرة من الجزيئات الضخمة والتي من المُمكن أن تُغير كيفية تصرف الجُزيئات بواسطة تزاحم الجزيئات الضخمة.[19]
على الرغم من أنهُ قد يُعتقد أنَّ العصارة الخلوية عبارة عن محلول بسيط من الجُزيئات، إلا أنها تمتلك مُستوياتٍ مُتعددة من التنظيم الخلوي،[20] والذي يتضمن تَدرُج تركيز الجزيئات الصغيرة مثل الكالسيوم والمركبات الإنزيمية الكبيرة التي تعمل معًا في المسارات الأيضية، ويتضمن أيضًا المركبات البروتينية مِثل الجسيمات البروتينية والجسميات الكربوكسيلية التي تُطَوِق وتفصل أجزاء العُصارة الخلوية.[21]
التعريف
استُعمِلَ مُصطلح العُصارة الخلوية (السيتوسول) للمرةِ الأولى عام 1965 بواسطة هنري لارد، وكانُ في البداية يُشير إلى السائل المُنتج من تكسيرِ الخلايا وتكوير جميع المُكونات غير القابلة للذوبان بواسطة المِنْبَذَة الفائِقَة،[22][23]، وبما أنَّ المُستخرج الخلوي القابِل للذوبان غير مُطابق للجزء السيتوبلازمي الخلوي القابل للذوبان فقد سُمي باسم الجزء السيتوبلازمي.[24]
يُستخدم مُصطلح العُصارة الخلوية حاليًا ليُشير إلى الجانِب السائل للسيتوبلازم في الخلية السليمة،[24] وبالتالي يُستثنى منها أي جزء سيتوبلازمي مُضمن داخل العُضيات الخلوية. نظرًا لاحتمال الخَلطِ بين استعمال كلمة العُصارة الخلوية للإشارة إلى مُستخرجات الخلايا والجزء القابل للذوبان من السيتوبلازم في الخلايا السليمة، فإنَّ عبارة "السيتوبلازم المائي" أصبحت تُستخدم لوصف المحتويات السائلة في سيتوبلازم الخلايا الحية.[23]
استُخدمت مُصطلحات أخرى قبل هذا المُصطلح لوصف السائل الخَلوي، ولكنها لا تُعتبر مترادفات؛ لأنَّ طبيعتها غير واضحة تمامًا، ومن الأمثلة عليها البروتوبلازم.[24]
الخواص والمكونات
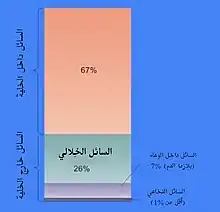
تَختلف نِسبة العصارة الخلوية من حجم الخلية، فمثلًا تُشكل العصارة الخلوية الجُزء الأكبر من بنية الخلية في البكتيريا،[25] أما في الخلايا النباتية فإنَّ الجزء الأكبر تحتله الفجوة العصارية المركزية الكبيرة.[26] تتكون العُصارة الخلوية في مُعظمها من الماء والأيونات المذابة والجزيئات الصغيرة والجزيئات الكبيرة القابلة للذوبان في الماء (مثل البروتينات)، كما أنَّ مُعظم هذه الجزيئات غير البروتينية تمتلك كتلة جزيئية أقل من 300 دالتون.[27] يُعتبر هذا الخليط من الجزيئات الصغيرة مُعقدًا للغاية؛ وذلك لأنَّ مجموعة مُتنوعة هائِلة من هذه الجُزيئات تُعتبر من نواتج عملية الأيض، فمثلًا أكثر من 200 ألف جُزيء صغير مُختلف قد يُصنع في النباتات، وذلك إن لم تكن جميع هذه الجُزيئات موجودة في نفس النوع أو في خلية واحدة.[28] تتوقع بعض التقديرات أنَّ عدد نواتج الأيض في الخلايا الوحيدة مثل الإشريكية القولونية وفطر الخميرة يبلغ أقل من 1000 ناتج.[29][30]
الماء
مُعظم العُصارة الخلوية ماء، حيثُ يُشكل حوالي 70% من الحجم الكُلي للخلية النموذجية.[17] الأس الهيدروجيني للسائل داخل الخلية هو 7.4،[31] بينما الأس الهيدروجيني للعُصارة الخلوية البشرية يترواح بين 7.0 - 7.4، وغالبًا ما يكون أعلى إذا كانت الخلية في طور النُمو.[32] إنَّ لزوجة السيتوبلازم هي تقريبًا نفس لزوجة الماء النقي، وعلى الرغم من هذا فإنَّ انتشار الجزيئات الصغيرة من خلال السيتوبلازم أبطأ بحوالي أربعة أضعاف مما هيَ عليه في الماء النقي؛ وذلك غالبًا بسبب التصادمات مع الأعداد الكبيرة من الجزيئات الضخمة في العصارة الخلوية.[33] بَحثت دراسات في الروبيان الملحي حولَ كيفية تأثير الماء على وظائف الخلية، حيثُ أظهرت أنَّ انخفاض 20% في كمية الماء في الخلية يؤدي إلى تثبيط عملية الأيض، وعندما يصل مُستوى الماء إلى أقل من 70% من المستوى الاعتيادي فإنَّ عملية الأيض تنخفض تدريجيًا وبالتالي سوفَ تجف الخلية وتتوقف جميع النشاطات الأيضية.[23]
على الرغم من أنَّ الماء مُهم للحياة، إلا أنَّ بنية الماء في العُصارة الخلوية غير مفهومة جيدًا؛ وذلك غالبًا لأنَّ الأساليب مثل مطيافية الرنين المغناطيسي النووي تُعطي معلوماتٍ فقط عن متوسطِ بنية الماء في العُصارة الخلوية، ولا تستطيع قياس الاختلافات الموضعية في المقياس المجهري.[18]
تعتمد وُجهة النظر التقليدية حول الماء في الخلايا أنَّ 5% من هذا الماء مُرتبطة بقوة في المذابات أو الجزيئات الضخمة كما يحدث في عملية التذاوب، في حين أنَّ غالبية الماء الأخرى تمتلك نفس بُنية الماء النقي.[23] ماء التذاوب غير نشط اسموزيًا وقد يمتلك خواص المُذيبات المُختلفة، بحيثُ تستثنى بعض الجُزيئات الذائبة، ويتركز بعضها الآخر.[34][35] على الرغم من ذلك، يرى آخرون أنَّ آثار التراكيز العالية من الجزيئات الضخمة في الخلايا يمتد في جميع أنحاء العُصارة الخلوية، وأنَّ الماء في الخلايا يتصرف بشكلٍ مُختلف عن الماء في المحاليل المُخففة،[36] وتتضمن هذه الآراء الاقتراح بأنَّ الخلايا تحتوي على مناطق ذات كثافة عالية ومنخفضة للماء، مما قد يؤثر على نطاقٍ واسع على هياكل ووظائف الأجزاء الأخرى من الخلية.[18][37] مع ذلك، فإنَّ الاستعمال المُتقدم المُباشر لمطيافية الرنين المغناطيسي النووي لقياس حركة الماء في الخلايا الحية يُظهر ما يُناقض هذه الفكرة، حيث يُشير أنَّ 85% من ماء الخلية يتصرف مثل الماء النقي، والجزء المُتبقي أقلُ حركةً ويُحتمل ارتباطه مع الجزيئات الضخمة.[38]
الأيونات
تختلف تراكيز الأيونات الأخرى في العُصارة الخلوية تمامًا عن تلك الموجودة في السائل خارج الخلية، وأيضًا تحتوي العُصارة الخلوية على كمياتٍ أكبر من الجزيئات الضخمة المَشحونة مثل البروتينات والأحماض النووية عن تلك الموجودة خارج بنية الخلية.
| الأيون | التركيز في العصارة الخلوية (ميلي مولار) | التركيز في الدم (ميلي مولار) |
|---|---|---|
| بوتاسيوم | 139 | 4 |
| صوديوم | 12 | 145 |
| كلوريد | 4 | 116 |
| بيكربونات | 12 | 29 |
| الأحماض الأمينية في البروتينات | 138 | 9 |
| مغنسيوم | 0.8 | 1.5 |
| كالسيوم | <0.0002 | 1.8 |
على عكسِ السائل خارج الخلية، فإنَّ العُصارة الخلوية تحتوي على تركيزٍ عالٍ من أيونات البوتاسيوم وتركيزٍ مُنخفض من أيونات الصوديوم،[40] وهذا الاختلاف مُهم جدًا في التنظيم الاسموزي؛ لأنه إذا كانت مُستويات الأيونات داخل الخلية كما في خارجها، فإنَّ الماء سوف يدخل اسموزيًا باستمرارٍ؛ وذلك لأنَّ مستويات الجزيئات الضخمة داخل الخلايا أعلى من مستوياتها خارجها. بدلًا من ذلك، فإنَّ أيونات الصوديوم تُقذف وأيونات البوتاسيوم تُؤخذ عبر مضخة الصوديوم والبوتاسيوم، ثم تتدفق أيونات البوتاسيوم بناءً على تدرج تركيزها عبر القنوات الأيونية الانتقائية للبوتاسيوم، ويؤدي هذا الفقدان في الشُحنة الموجبة إلى حدوث الجهد الغشائي السالب، ولتحقيق التوازن في فرق الجهد، فإنَّ أيونات الكلوريد السالبة تخرج أيضًا من الخلية عبر قنوات الكلوريد الانتقائية، ويؤدي فقدان أيونات الصوديوم والكلوريد إلى التعويض عن التأثير الاسموزي للتركيز العالي للجزيئات العضوية داخل الخلية.[40]
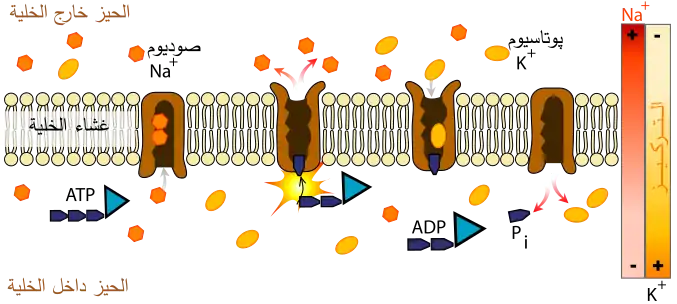
تستطيع الخلايا التَعامل مع التغيرات الاسموزية الكبيرة من خلال تجميعِ الحاميات الأسموزية مثل البيتائين والطرهالوز في عصارتها الخلوية.[40] تسمح بعض هذه الجزيئات للخلايا للبقاء على قيدِ الحياة ومنع تجفيفها تمامًا، كما تسمحُ للكائن الحي بالدخول في حالةٍ حيويةٍ مُعلقة أو موقوفة تُسمى الحيوية الخفية،[41] وفي هذه الحالة تُصبح العُصارة الخلوية والحاميات الأسموزية صلبةً كالزُجاج، حيث تساعد على تثبيت البروتينات والأغشية الخلوية من الآثار الضارة الناجمة عن الجفاف.[42]
يسمح تركيز الكاسيوم المُنخفض في العصارة الخلوية لأيونات الكالسيوم للعمل كمرسال ثاني في تأشير الكالسيوم، حيث تقوم هنا إشارة مثل هرمون أو جهد فعل بفتحِ قنوات الكالسيوم مما يسمح بتدفقِ الكالسيوم إلى العصارة الخلوية،[43] وتؤدي هذه الزيادة المُفاجئة في الكالسيوم العصاري الخلوي إلى تنشيط جزيئات الإشارة الأخرى مثل الكالموديولين وكيناز البروتين C.[44] قد تمتلك الأيونات الأخرى مثل الكلوريد والبوتاسيوم وظائف إشارية في العُصارة الخلوية، ولكن هذه الأيونات غير مفهومة جيدًا.[45]
الجزيئات الضخمة
تَذوب جزيئات البروتين التي لا ترتبط بالأغشية الخلوية أو بالهيكل الخلوي في العُصارة الخلوية. كمية البروتين في الخلايا عالية للغاية، حيثُ تقترب من 200 ملغ/مل، وتأخذ حوالي 20-30% من حجم العُصارة الخلوية،[19] وبالرغم من هذا، فإنهُ من الصعب القياس الدقيق لكمية البروتين الذائب في العُصارة الخلوية للخلايا السليمة؛ وذلك لأنَّ بعض البروتينات ترتبط بشكلٍ ضعيف مع الأغشية أو العضيات في الخلايا السليمة، كما أنها تنطلق نحوَ المحلول أثناء انحلال الخلايا.[23] عندما مُزقَ غشاء الخلايا البلازمي بحرص أثناء التجارب باستعمال الصابونين ودون الإضرار بأغشية الخلايا الأخرى، فإنَّ حوالي رُبع البروتين فقط انطلقَ نحو المحلول، كما كانت هذه الخلايا قادرة على تَخليق البروتينات في حالِ مُنحت أدينوسين ثلاثي الفوسفات وأحماض أمينية، وهذا يُدلل على أنَّ العديد من الإنزيمات تكون مُرتبطة بالهيكل الخلوي في العُصارة الخلوية.[46] مع ذلك، فإنَّه من غير المرجح صِحة فكرة أنَّ مُعظم البروتينات في الخلايا ترتبط بقوة في شبكة تُسمى الشبيكة الحُوَيْجِزية الدقيقة.[47]
تحتوي العُصارة الخلوية في بدائيات النوى على المجموع المورثي الخلوي، وذلك ضمنَ هيكلٍ يُسمى النوواني،[16] وهو عبارة عن كتلة غير منتظمة من الحمض النووي الريبوزي منقوص الأكسجين والمرتبط مع بروتينات تتحكم في نسخ وتضاعف الكروموسوم والبلازميد البكتيري. أما في حقيقات النوى، فإنَّ المجموع المورثي يتواجد في نواة الخلية، والتي تُعتبر مفصولة عن العُصارة الخلوية بواسطة المسام النووية التي تمنع انتشار أي جزيء قطره أكبر من 10 نانومتر.[15]
يؤدي التركيز العالي للجزيئات الضخمة في العُصارة الخلوية إلى تأثيرٍ يُسمى تزاحم الجزيئات الضخمة، والذي يحدث عند زيادة التركيز الفعال للجزيئات الضخمة الأخرى؛ وذلك لأنَّ الجزيئات الضخمة تمتلك حجمًا أقل للتحرك في العصارة الخلوية. قد يُنتِج هذا التزاحُم تغيراتٍ كبيرة في سرعة التفاعلات وحالة التوازن الكيميائي للتفاعلات في العصارة الخلوية.[19] من المهم بشكلٍ خاص قدرتها على تغيير ثوابت الانحلال من خلال تفضيل الارتباط مع الجزيئات الضخمة، وذلك كما يحدث عندما تتجمع بروتينات متعددة لتشكيل مركبات بروتينية، أو عندما يرتبط البروتين المرتبط بالحمض النووي الريبوزي منقوص الأكسجين مع المنطقة الهدف في المجموع المورثي.[48]
تنظيم العُصارة
بالرغم من أنَّ مُكونات العصارة الخلوية غير مُقسمة إلى مناطق بواسطة الأغشية الخلوية، إلا أنَّ هذه المكونات لا تختلط بشكلٍ عشوائي، وعلاوةً على ذلك، فإنهُ توجد عدة مستويات تنظيمية تُساعد على تموضُع الجزيئات لتحديد مواقعها ضمن العصارة الخلوية.[20]
تدرجات التركيز
على الرغم من أنَّ الجزيئات الصَغيرة تنتشر بسرعة في العُصارة الخلوية، إلا أنهُ قد تحدث تدرجات في التركيز ضمن العُصارة نفسها، ومن الأمثلة المَدروسة على ذلك "شرارات الكالسيوم" التي تُنتج لفترةٍ قصيرة حول منطقة قناة الكالسيوم المفتوحة،[49] والتي يبلغ قطرها 2 ميكرومتر وتستمر فقط لبضعة أجزاء من الثانية، وأيضًا قد تندمج العديد من الشرارات لتُشكِلَ تدرجاتٍ أكبر في التركيز، وتُسمى في هذه الحالة "موجات الكالسيوم".[50] قد تنتج تدرجات في التركيز في الجزيئات الصغيرة الأخرى مثل الأكسجين والأدينوسين ثلاثي الفوسفات، حيث قد تنتج التدرجات في الخلايا حول تجمعات الميتوكندريا، وهذه التدرجات أيضًا غير مفهومة جيدًا.[51][52]
المركبات البروتينية
يُمكن أن ترتبط البروتينات لتكوين مركبات بروتينية، والتي غالبًا ما تحتوي على مجموعة من البروتينات متماثلة الوظائف، كما هو الحال في الإنزيمات التي تقوم بعدة خطوات في نفس المسار الأيضي.[53] يُساعد هذا التنظيم البروتيني في توجيه الركيزة، الذي يسمح بمرور مُنتج أحد الإنزيمات مباشرةً إلى الإنزيم التالي في المسار دون الخروج إلى المحلول،[54] وأيضًا يُساعد التوجيه في زيادة فعالية وسرعة حدوث المسار الأيضي، بدلًا من توزع الإنزيمات عشوائيًا في العُصارة الخلوية، كما يُساعد التوجيه في تجنب انطلاق أي مركبات وسطية غير مستقرة.[55] على الرغم من أنَّ مجموعة واسعة من المسارات الأيضية تتضمن إنزيمات ترتبط بقوة مع بعضها البعض، إلا أنَّ مساراتٍ أخرى قد تتضمن مركبات ترتبط بشكلٍ رخو،[56][57] وبالتالي يصعب دراستها خارج الخلية؛ لذلك أهمية هذه المركبات في عملية الأيض لا تزال غير واضحة بشكلٍ عام.
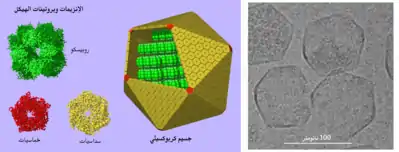
الحيز البروتيني
تحتوي بعض المركبات البروتينية على تجويف مركزي كبير معزول عن باقي العُصارة الخلوية، ومن الأمثلة عليه الحيز المُغلق في الجسيم البروتيني،[58] حيثُ تشكل مجموعة من الوحدات الفرعية تجويف يُشبه المخزن يحتوي على البروتياز الذي يعمل على تحليل بروتينات العصارة الخلوية، وبما أنَّ هذه العملية قد تكون ضارة إذا اختلطت مع باقي العُصارة الخلوية، فإنَّ المخزن يُحاط بمجموعة من البروتينات التنظيمية التي تتعرف على البروتينات مع توجيهها إلى منطقة التحلل (واسِم اليوبيكويتين) وأيضًا إدخالها في التجويف المحلل للبروتين.[59]
هُناك مجموعة أخرى كبيرة من الحيز البروتيني تكون عبارة عن حيز بكتيري دقيق، والتي تتكون من هيكل بروتيني يُغلف مجموعة من الإنزيمات المختلفة،[60] ويكون قطر هذا الحيز حوالي 100-200 نانومتر ويتكون من بروتينات متشابكة.[61] تُعتبر الجسيمات الكربوكسيلية من الأمثلة المفهومة جيدًا حول هذا الموضوع، حيث تحتوي على إنزيمات مُضمنة في تثبيت الكربون مثل روبيسكو.[21]
هيكل الخلية
على الرغم من أنَّ الهيكل الخلوي ليسَ جزءً من العُصارة الخلوية، إلا أنَّ وجود هذه الشبكة من الخيوط يُقيد انتشار الجزيئات الكبيرة في الخلية، مثلًا، في العديد من الدراسات استبعدت الجزيئات الاستشفافية (المتتبّعة)[lower-alpha 2] الأكبر من 25 نانومتر (تقريبًا نفس حجم الريبوسوم)[62] من جزء من العصارة الخلوية حول حواف الخلية وبجانب النواة.[63][64] قد يحتوي الحيز المُستبعدة منه هذه الجزيئات على شبكة من ألياف الأكتين أكثر كثافة من الجزء المتبقي من العصارة الخلوية. يمكن أن تؤثر هذه النطاقات الصغيرة على توزيع الهياكل الكبيرة مثل الرايبوسومات والعضيات ضمن العصارة الخلوية، وذلك من خلال إبعادها من بعض المناطق وتركيزها في مناطق أخرى.[65]
الوظيفة
تتعدد وظائف العُصارة الخلوية، حيث تعتبر موقع حدوث العديد من عمليات الخلية، ومن الأمثلة على هذه العمليات توصيل الإشارة من غشاء الخلية إلى مواقع مُعينة داخل الخلية مثل النواة[66] أو العضيات.[67] أيضًا تعتبر العصارة الخلوية موقعًا للعديد من عمليات الانقسام السيتوبلازمي بعد تفكك الغشاء النووي في الانقسام المتساوي.[68] من الوظائف الرئيسية الأخرى للعصارة الخلوية، نقل المُنتجات الأيضية من مواقع الإنتاج إلى مواقع الاستخدام، وهذا الأمر بسيط نسبيًا بالنسبة للجزيئات القابلة للذوبان في الماء، مثل الأحماض الأمينية التي تستطيع الانتشار بسرعة من خلال العصارة الخلوية، ومع ذلك فإنَّ الجزيئات الكارهة للماء مثل الأحماض الدهنية والستيرولات، يُمكن أن تنتقل عبر العصارة الخلوية بواسطة بروتينات ربط خاصة، والتي تنقل هذه الجزيئات بين أغشية الخلية.[69][70] يُمكن أيضًا أن تنتقل الجزيئات المأخوذة بواسطة الإدخال الخلوي أو التي في طريقها إلى أن تفرز من خلال العصارة الخلوية وعبر الحويصلات[71] التي هي عبارة عن كريات صغيرة من الدهون والتي تنتقل على طول الهيكل الخلوي بواسطة بروتينات حركية.[72]
العصارة الخلوية هي موقع معظم عمليات الأيض لبدائيات النوى،[25] كما أنَّ نسبة كبيرة من عمليات الأيض في حقيقات النوى تحدث فيها. مثلًا، في الثدييات تتواجد حوالي نصف بروتينات الخلية في العصارة الخلوية.[73] تتوفر بيانات أكثر حول الموضوع في الخميرة، حيث تشير عمليات الاستبناء الأيضي أنَّ غالبية العمليات الأيضية ونواتج الأيض تحدث في العصارة الخلوية.[74] غالبية المسارات الأيضية التي تحدث في العصارة الخلوية في الحيوانات هي عبارة عن اصطناع حيوي للبروتين ومسار فوسفات البنتوز وتحلل السكر واستحداث الجلوكوز.[75] قد تكون أماكن حدوث المسارات الأيضية مُختلفة في الكائنات الحية الأخرى، مثلًا يحدث تصنيف الأحماض الدهنية في البلاستيدات الخضراء في النباتات،[76][77] وفي صانعات القمة في معقدات القمة.[78]
العصارة الخلوية مقابل السيتوبلازم
غالبًا ما يُخلط بين العصارة الخلوية والسيتوبلازم، ولكنِهما يختلفان تمامًا في الخلايا الحية، حيث يتكون السيتوبلازم من جميع المحتويات داخل الخلية باستثناء نواة الخلية، أما العصارة الخلوية فهي فقط الجزء السائل أو المائي من السيتوبلازم. أي أنَّ السيتوبلازم هو عبارة عن كُل المساحة الموجودة بين النواة وغشاء الخلية، ويتكون من العُصارة الخلوية والعضيات الأخرى، وتعتبر العصارة الخلوية من أهم المواد في السيتوبلازم.[79]
التسمية
تُنطق كلمة Cytosol (سيتوسول) حَسب الألفبائية الصوتية الدولية (/ˈsaɪtəˌsɒl/)، وحسب قاموس ميريام وبستر، فإنَّ كلمة Cytosol (سيتوسول) عُرفت لأول مرة في عام 1965.[80]
حسب قاموس التراث الأمريكي للغة الإنجليزية، فإنَّ كلمة Cytosol (سيتوسول) تتكون من مقطعين، الأول cyto- وهي سابقة بمعنى الخلية أو خلوي، والثاني sol وهو اختصار لكلمة solution والتي تعني محلول.[81]
ملاحظات
المراجع
- معرف أنطولوجية الجينات: http://amigo.geneontology.org/amigo/term/GO:0005829 — تاريخ الاطلاع: 12 أكتوبر 2019 — الاصدار 2019-10-07
- Goodsell DS (June 1991). "Inside a living cell". Trends Biochem. Sci. 16 (6): 203–6. doi:10.1016/0968-0004(91)90083-8. PMID 1891800. الوسيط
|CitationClass=تم تجاهله (مساعدة) - يُوسف حِتّي; أحمَد شفيق الخَطيب (2008). قامُوس حِتّي الطِبي للجَيب. بيروت، لبنان: مكتبة لبنان ناشرون. صفحة 112. ISBN 995310235X. الوسيط
|CitationClass=تم تجاهله (مساعدة);|access-date=بحاجة لـ|url=(مساعدة) - طَلال أبو غَزالة (2010). معجم طلال أبوغزاله لبراءات الاختراع. المناهل. صفحة 59. ISBN 9796500078502. الوسيط
|CitationClass=تم تجاهله (مساعدة);|access-date=بحاجة لـ|url=(مساعدة) - "معاني كلمة (cytosol) في موقع القاموس/طب". موقع القاموس. مؤرشف من الأصل في 21 يونيو 2018. اطلع عليه بتاريخ 27 يناير 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "معاني كلمة (cytosol) في موقع المعاني/طب". موقع قاموس المعاني. مؤرشف من الأصل في 10 يناير 2020. اطلع عليه بتاريخ 27 يناير 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "معاني كلمة (cytosol) في المعجم الطبي الموحد". المعجم الطبي الموحد. مؤرشف من الأصل في 17 أبريل 2019. اطلع عليه بتاريخ 27 يناير 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "الداء السيستيني- (العصارة الخلوية - Cytosol)". ويب طب. مؤرشف من الأصل في 1 يوليو 2017. اطلع عليه بتاريخ 9 فبراير 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - جابر القحطاني (2009). موسوعة جابِر لِطب الأعشاب: الجزء الأول. مكتبة العبيكان. صفحة 48. ISBN 9960540375. الوسيط
|CitationClass=تم تجاهله (مساعدة);|access-date=بحاجة لـ|url=(مساعدة) - عبد المنعم محمد الأعسر (2011). أُسس الكيمياء الحيوية. القاهرة، مصر: المكتبة الأكاديمية. صفحة 445. الوسيط
|CitationClass=تم تجاهله (مساعدة);|access-date=بحاجة لـ|url=(مساعدة) - عماد الدين وصفي (1998). عماد فسيولوجي النبات. القاهرة، مصر: المكتبة الأكاديمية. صفحة 8. الوسيط
|CitationClass=تم تجاهله (مساعدة);|access-date=بحاجة لـ|url=(مساعدة) - "ترجمة ومعنى matrix في قاموس المعاني". www.almaany.com. مؤرشف من الأصل في 21 يونيو 2018. اطلع عليه بتاريخ 27 يناير 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "ترجمة ومعنى cytoplasmic matrix في القاموس/طب". www.alqamoos.org. مؤرشف من الأصل في 21 يونيو 2018. اطلع عليه بتاريخ 27 يناير 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - LESLIE P. GARTNER; JAMES L. HIATT (2011). Concise Histology [المختصر في علم الأنسجة] (باللغة الإنجليزية). إلزيفير. صفحة 8. ISBN 9780702031144. الوسيط
|CitationClass=تم تجاهله (مساعدة);|access-date=بحاجة لـ|url=(مساعدة) - Peters R (2006). "Introduction to nucleocytoplasmic transport: molecules and mechanisms". Methods Mol. Biol. 322: 235–58. doi:10.1007/978-1-59745-000-3_17. ISBN 978-1-58829-362-6. PMID 16739728. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Thanbichler M, Wang S, Shapiro L (2005). "The bacterial nucleoid: a highly organized and dynamic structure". J Cell Biochem. 96 (3): 506–21. doi:10.1002/jcb.20519. PMID 15988757. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Luby-Phelps K (2000). "Cytoarchitecture and physical properties of cytoplasm: volume, viscosity, diffusion, intracellular surface area" (PDF). Int. Rev. Cytol. 192: 189–221. doi:10.1016/S0074-7696(08)60527-6. ISBN 978-0-12-364596-8. PMID 10553280. مؤرشف من الأصل (PDF) في 19 يوليو 2011. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wiggins PM (1 December 1990). "Role of water in some biological processes". Microbiol. Rev. 54 (4): 432–49. PMC 372788. PMID 2087221. مؤرشف من الأصل في 11 يونيو 2011. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ellis RJ (أكتوبر 2001). "Macromolecular crowding: obvious but underappreciated". Trends Biochem. Sci. 26 (10): 597–604. doi:10.1016/S0968-0004(01)01938-7. PMID 11590012. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Norris V, den Blaauwen T, Cabin-Flaman A (March 2007). "Functional Taxonomy of Bacterial Hyperstructures". Microbiol. Mol. Biol. Rev. 71 (1): 230–53. doi:10.1128/MMBR.00035-06. PMC 1847379. PMID 17347523. مؤرشف من الأصل في 14 يوليو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Badger MR, Price GD (February 2003). "CO2 concentrating mechanisms in cyanobacteria: molecular components, their diversity and evolution". J. Exp. Bot. 54 (383): 609–22. doi:10.1093/jxb/erg076. PMID 12554704. مؤرشف من الأصل في 25 يوليو 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lardy, H. A. 1965. On the direction of pyridine nucleotide oxidation-reduction reactions in gluconeogenesis and lipogenesis. In: Control of energy metabolism, edited by B. Chance, R. Estabrook, and J. R. Williamson. New York: Academic, 1965, p. 245, . نسخة محفوظة 18 ديسمبر 2016 على موقع واي باك مشين.
- Clegg JS (February 1984). "Properties and metabolism of the aqueous cytoplasm and its boundaries". Am. J. Physiol. 246 (2 Pt 2): R133–51. PMID 6364846. مؤرشف من الأصل في 26 مارس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cammack, Richard; Teresa Atwood; Attwood, Teresa K.; Campbell, Peter Scott; Parish, Howard I.; Smith, Tony; Vella, Frank; Stirling, John (2006). Oxford dictionary of biochemistry and molecular biology. Oxford [Oxfordshire]: Oxford University Press. ISBN 0-19-852917-1. OCLC 225587597. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Hoppert M, Mayer F (1999). "Principles of macromolecular organization and cell function in bacteria and archaea". Cell Biochem. Biophys. 31 (3): 247–84. doi:10.1007/BF02738242. PMID 10736750. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bowsher CG, Tobin AK (April 2001). "Compartmentation of metabolism within mitochondria and plastids". J. Exp. Bot. 52 (356): 513–27. doi:10.1093/jexbot/52.356.513. PMID 11373301. مؤرشف من الأصل في 15 فبراير 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Goodacre R, Vaidyanathan S, Dunn WB, Harrigan GG, Kell DB (May 2004). "Metabolomics by numbers: acquiring and understanding global metabolite data" (PDF). Trends Biotechnol. 22 (5): 245–52. doi:10.1016/j.tibtech.2004.03.007. PMID 15109811. مؤرشف من الأصل (PDF) في 17 ديسمبر 2008. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Weckwerth W (2003). "Metabolomics in systems biology". Annu Rev Plant Biol. 54: 669–89. doi:10.1146/annurev.arplant.54.031902.135014. PMID 14503007. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Reed JL, Vo TD, Schilling CH, Palsson BO (2003). "An expanded genome-scale model of Escherichia coli K-12 (iJR904 GSM/GPR)". Genome Biol. 4 (9): R54. doi:10.1186/gb-2003-4-9-r54. PMC 193654. PMID 12952533. مؤرشف من الأصل في 11 يناير 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Förster J, Famili I, Fu P, Palsson BØ, Nielsen J (February 2003). "Genome-Scale Reconstruction of the Saccharomyces cerevisiae Metabolic Network". Genome Res. 13 (2): 244–53. doi:10.1101/gr.234503. PMC 420374. PMID 12566402. مؤرشف من الأصل في 26 مارس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Roos A, Boron WF (April 1981). "Intracellular pH". Physiol. Rev. 61 (2): 296–434. PMID 7012859. مؤرشف من الأصل في 26 مايو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bright, G R; Fisher, GW; Rogowska, J; Taylor, DL (1987). "Fluorescence ratio imaging microscopy: temporal and spatial measurements of cytoplasmic pH". The Journal of Cell Biology. 104 (4): 1019–1033. doi:10.1083/jcb.104.4.1019. PMC 2114443. PMID 3558476. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Verkman AS (January 2002). "Solute and macromolecule diffusion in cellular aqueous compartments". Trends Biochem. Sci. 27 (1): 27–33. doi:10.1016/S0968-0004(01)02003-5. PMID 11796221. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Fulton AB (September 1982). "How crowded is the cytoplasm?". Cell. 30 (2): 345–7. doi:10.1016/0092-8674(82)90231-8. PMID 6754085. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Garlid KD (2000). "The state of water in biological systems". Int. Rev. Cytol. 192: 281–302. doi:10.1016/S0074-7696(08)60530-6. ISBN 978-0-12-364596-8. PMID 10553283. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Chaplin M (November 2006). "Do we underestimate the importance of water in cell biology?". Nat. Rev. Mol. Cell Biol. 7 (11): 861–6. doi:10.1038/nrm2021. PMID 16955076. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wiggins PM (June 1996). "High and low density water and resting, active and transformed cells". Cell Biol. Int. 20 (6): 429–35. doi:10.1006/cbir.1996.0054. PMID 8963257. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Persson E, Halle B (April 2008). "Cell water dynamics on multiple time scales". Proc. Natl. Acad. Sci. U.S.A. 105 (17): 6266–71. doi:10.1073/pnas.0709585105. PMC 2359779. PMID 18436650. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lodish, Harvey F. (1999). Molecular cell biology. New York: Scientific American Books. ISBN 0-7167-3136-3. OCLC 174431482. مؤرشف من الأصل في 16 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lang F (October 2007). "Mechanisms and significance of cell volume regulation". J Am Coll Nutr. 26 (5 Suppl): 613S–623S. doi:10.1080/07315724.2007.10719667. PMID 17921474. مؤرشف من الأصل في 10 يناير 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Sussich F, Skopec C, Brady J, Cesàro A (August 2001). "Reversible dehydration of trehalose and anhydrobiosis: from solution state to an exotic crystal?". Carbohydr. Res. 334 (3): 165–76. doi:10.1016/S0008-6215(01)00189-6. PMID 11513823. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Crowe JH, Carpenter JF, Crowe LM (1998). "The role of vitrification in anhydrobiosis". Annu. Rev. Physiol. 60: 73–103. doi:10.1146/annurev.physiol.60.1.73. PMID 9558455. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Berridge MJ (1 March 1997). "Elementary and global aspects of calcium signalling". J. Physiol. 499 (Pt 2): 291–306. doi:10.1113/jphysiol.1997.sp021927. PMC 1159305. PMID 9080360. مؤرشف من الأصل في 26 مايو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Kikkawa U, Kishimoto A, Nishizuka Y (1989). "The protein kinase C family: heterogeneity and its implications". Annu. Rev. Biochem. 58: 31–44. doi:10.1146/annurev.bi.58.070189.000335. PMID 2549852. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Orlov SN, Hamet P (April 2006). "Intracellular monovalent ions as second messengers". J. Membr. Biol. 210 (3): 161–72. doi:10.1007/s00232-006-0857-9. PMID 16909338. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Hudder A, Nathanson L, Deutscher MP (ديسمبر 2003). "Organization of Mammalian Cytoplasm". Mol. Cell. Biol. 23 (24): 9318–26. doi:10.1128/MCB.23.24.9318-9326.2003. PMC 309675. PMID 14645541. مؤرشف من الأصل في 7 مارس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Heuser J (2002). "Whatever happened to the 'microtrabecular concept'?". Biol Cell. 94 (9): 561–96. doi:10.1016/S0248-4900(02)00013-8. PMID 12732437. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Zhou HX, Rivas G, Minton AP (2008). "Macromolecular crowding and confinement: biochemical, biophysical, and potential physiological consequences". Annu Rev Biophys. 37: 375–97. doi:10.1146/annurev.biophys.37.032807.125817. PMC 2826134. PMID 18573087. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wang SQ, Wei C, Zhao G (April 2004). "Imaging microdomain Ca2+ in muscle cells". Circ. Res. 94 (8): 1011–22. doi:10.1161/01.RES.0000125883.68447.A1. PMID 15117829. مؤرشف من الأصل في 6 يوليو 2010. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Jaffe LF (November 1993). "Classes and mechanisms of calcium waves". Cell Calcium. 14 (10): 736–45. doi:10.1016/0143-4160(93)90099-R. PMID 8131190. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Aw, T.Y. (2000). "Intracellular compartmentation of organelles and gradients of low molecular weight species". Int Rev Cytol. 192: 223–53. doi:10.1016/S0074-7696(08)60528-8. ISBN 978-0-12-364596-8. PMID 10553281. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Weiss JN, Korge P (20 July 2001). "The cytoplasm: no longer a well-mixed bag". Circ. Res. 89 (2): 108–10. PMID 11463714. مؤرشف من الأصل في 3 فبراير 2009. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Srere PA (1987). "Complexes of sequential metabolic enzymes". Annu. Rev. Biochem. 56: 89–124. doi:10.1146/annurev.bi.56.070187.000513. PMID 2441660. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Perham RN (2000). "Swinging arms and swinging domains in multifunctional enzymes: catalytic machines for multistep reactions". Annu. Rev. Biochem. 69: 961–1004. doi:10.1146/annurev.biochem.69.1.961. PMID 10966480. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Huang X, Holden HM, Raushel FM (2001). "Channeling of substrates and intermediates in enzyme-catalyzed reactions". Annu. Rev. Biochem. 70: 149–80. doi:10.1146/annurev.biochem.70.1.149. PMID 11395405. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Mowbray J, Moses V (June 1976). "The tentative identification in Escherichia coli of a multienzyme complex with glycolytic activity". Eur. J. Biochem. 66 (1): 25–36. doi:10.1111/j.1432-1033.1976.tb10421.x. PMID 133800. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Srivastava DK, Bernhard SA (November 1986). "Metabolite transfer via enzyme-enzyme complexes". Science. 234 (4780): 1081–6. doi:10.1126/science.3775377. PMID 3775377. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Groll M, Clausen T (December 2003). "Molecular shredders: how proteasomes fulfill their role". Curr. Opin. Struct. Biol. 13 (6): 665–73. doi:10.1016/j.sbi.2003.10.005. PMID 14675543. مؤرشف من الأصل في 6 يونيو 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Nandi D, Tahiliani P, Kumar A, Chandu D (March 2006). "The ubiquitin-proteasome system" (PDF). J. Biosci. 31 (1): 137–55. doi:10.1007/BF02705243. PMID 16595883. مؤرشف من الأصل (PDF) في 11 أغسطس 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bobik, T. A. (2007). "Bacterial Microcompartments" (PDF). Microbe. Am Soc Microbiol. 2: 25–31. مؤرشف من الأصل (PDF) في 02 أغسطس 2008. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Yeates TO, Kerfeld CA, Heinhorst S, Cannon GC, Shively JM (August 2008). "Protein-based organelles in bacteria: carboxysomes and related microcompartments". Nat. Rev. Microbiol. 6 (9): 681–691. doi:10.1038/nrmicro1913. PMID 18679172. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cate JH (November 2001). "Construction of low-resolution x-ray crystallographic electron density maps of the ribosome". Methods. 25 (3): 303–8. doi:10.1006/meth.2001.1242. PMID 11860284. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Provance DW, McDowall A, Marko M, Luby-Phelps K (1 October 1993). "Cytoarchitecture of size-excluding compartments in living cells". J. Cell Sci. 106 (2): 565–77. PMID 7980739. مؤرشف من الأصل في 26 مارس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Luby-Phelps K, Castle PE, Taylor DL, Lanni F (July 1987). "Hindered diffusion of inert tracer particles in the cytoplasm of mouse 3T3 cells". Proc. Natl. Acad. Sci. U.S.A. 84 (14): 4910–3. doi:10.1073/pnas.84.14.4910. PMC 305216. PMID 3474634. مؤرشف من الأصل في 26 مايو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Luby-Phelps K (June 1993). "Effect of cytoarchitecture on the transport and localization of protein synthetic machinery". J. Cell. Biochem. 52 (2): 140–7. doi:10.1002/jcb.240520205. PMID 8366131. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Kholodenko BN (June 2003). "Four-dimensional organization of protein kinase signaling cascades: the roles of diffusion, endocytosis and molecular motors". J. Exp. Biol. 206 (Pt 12): 2073–82. doi:10.1242/jeb.00298. PMID 12756289. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Pesaresi P, Schneider A, Kleine T, Leister D (December 2007). "Interorganellar communication". Curr. Opin. Plant Biol. 10 (6): 600–6. doi:10.1016/j.pbi.2007.07.007. PMID 17719262. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Winey M, Mamay CL, O'Toole ET (June 1995). "Three-dimensional ultrastructural analysis of the Saccharomyces cerevisiae mitotic spindle". J. Cell Biol. 129 (6): 1601–15. doi:10.1083/jcb.129.6.1601. PMC 2291174. PMID 7790357. مؤرشف من الأصل في 24 أغسطس 2005. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Weisiger RA (October 2002). "Cytosolic fatty acid binding proteins catalyze two distinct steps in intracellular transport of their ligands". Mol. Cell. Biochem. 239 (1–2): 35–43. doi:10.1023/A:1020550405578. PMID 12479566. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Maxfield FR, Mondal M (June 2006). "Sterol and lipid trafficking in mammalian cells". Biochem. Soc. Trans. 34 (Pt 3): 335–9. doi:10.1042/BST0340335. PMID 16709155. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Pelham HR (August 1999). "The Croonian Lecture 1999. Intracellular membrane traffic: getting proteins sorted". Philos. Trans. R. Soc. Lond. B Biol. Sci. 354 (1388): 1471–8. doi:10.1098/rstb.1999.0491. PMC 1692657. PMID 10515003. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Kamal A, Goldstein LS (February 2002). "Principles of cargo attachment to cytoplasmic motor proteins". Curr. Opin. Cell Biol. 14 (1): 63–8. doi:10.1016/S0955-0674(01)00295-2. PMID 11792546. مؤرشف من الأصل في 12 فبراير 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Foster LJ, de Hoog CL, Zhang Y (April 2006). "A mammalian organelle map by protein correlation profiling". Cell. 125 (1): 187–99. doi:10.1016/j.cell.2006.03.022. PMID 16615899. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Herrgård, MJ; Swainston, N; Dobson, P; Dunn, WB; Arga, KY; Arvas, M; Blüthgen, N; Borger, S; Costenoble, R; et al. (October 2008). "A consensus yeast metabolic network reconstruction obtained from a community approach to systems biology". Nature Biotechnology. 26 (10): 1155–60. doi:10.1038/nbt1492. PMC 4018421. PMID 18846089. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Stryer, Lubert; Berg, Jeremy Mark; Tymoczko, John L. (2002). Biochemistry. San Francisco: W.H. Freeman. ISBN 0-7167-4684-0. OCLC 179705944. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ohlrogge J, Pollard M, Bao X (December 2000). "Fatty acid synthesis: from CO2 to functional genomics". Biochem. Soc. Trans. 28 (6): 567–73. doi:10.1042/BST0280567. PMID 11171129. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ohlrogge JB, Kuhn DN, Stumpf PK (March 1979). "Subcellular localization of acyl carrier protein in leaf protoplasts of Spinacia oleracea". Proc. Natl. Acad. Sci. U.S.A. 76 (3): 1194–8. doi:10.1073/pnas.76.3.1194. PMC 383216. PMID 286305. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Goodman CD, McFadden GI (January 2007). "Fatty acid biosynthesis as a drug target in apicomplexan parasites". Curr Drug Targets. 8 (1): 15–30. doi:10.2174/138945007779315579. PMID 17266528. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Cytosol vs Cytoplasm". 19 ديسمبر 2017. مؤرشف من الأصل في 15 أغسطس 2018. اطلع عليه بتاريخ 30 يناير 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Definition of CYTOSOL". ميريام وبستر. مؤرشف من الأصل في 2 يونيو 2017. اطلع عليه بتاريخ 30 يناير 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Wordnik-Etymologies of Cytosol word". Wordnik.com. مؤرشف من الأصل في 30 يناير 2018. اطلع عليه بتاريخ 30 يناير 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة)
كتب
- Wheatley, Denys N.; Pollack, Gerald H.; Cameron, Ivan L. (2006). Water and the Cell [الماء والخلية] (باللغة الإنجليزية). Berlin: Springer. ISBN 1-4020-4926-9. OCLC 71298997. الوسيط
|CitationClass=تم تجاهله (مساعدة) - LESLIE P. GARTNER; JAMES L. HIATT (2011). Concise Histology [المختصر في علم الأنسجة] (باللغة الإنجليزية). Philadelphia: إلزيفير. ISBN 9780702031144. الوسيط
|CitationClass=تم تجاهله (مساعدة);|access-date=بحاجة لـ|url=(مساعدة) - John McBrewster; Agnes F. Vandome; Frederic P. Miller (2009). Cytosol [السيتوسول] (باللغة الإنجليزية). Saarbrücken: VDM Publishing. ISBN 9786130260729. الوسيط
|CitationClass=تم تجاهله (مساعدة);|access-date=بحاجة لـ|url=(مساعدة)
 بوابة تشريح
بوابة تشريح- بوابة علم الأحياء الخلوي والجزيئي
