شبكة إندوبلازمية
الشبكة الهيولية الباطنة[1] أو الداخلية[2] أو الشبكة الإندوبلازمية[3] أو الشبكة البلازمية الداخلية[4] (بالإنجليزية: Endoplasmic reticulum) هي عضية خلوية تتواجد في خلايا حقيقيات النوى وتتكون من شبكة مترابطة من النبيبات والحويصلات و الأكياس الغشائية المسطحة أو بنيات تشبه الأنابيب تسمى صهاريج. ترتبط في مناطق معينة بالغشاء البلازمي ومع الغشاء النووي في مناطق أخرى ،
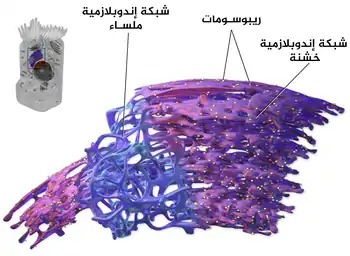

الأغشية في الشبكة الإندوبلازمية امتداد للغشاء النووي الخارجي، وتوجد الشبكة الإندوبلازمية في معظم أنواع خلايا حقيقيات النوى لكنها غائبة في الخلايا الدموية الحمراء والخلايا المنوية.
يوجد نوعان من الشبكة الإندوبلازمية: الشبكة الإندوبلازمية الخشنة والشبكة الإندوبلازمية الملساء، الوجه الخارجي (السيتوسولي) للشبكة الخشنة مرصّع بالريبوسومات التي هي مواقع تخليق البروتين. الشبكة الخشنة بارزة ووفيرة في الخلايا بشكل أكبر خاصة خلايا الكبد. الشبكة الملساء لا تملك ريبوسومات على سطحها وهي قليلة في معظم الخلايا وتعمل في تخليق الليبيدات وإنتاج الستيرويدات والهرمونات وإزالة سمية النواتج الطبيعية للأيض والكحول والأدوية، وهي وفيرة في خلايا الخصية والمبيض والغدة الدهنية.
تقوم الشبكة الإندوبلازمية بالعديد من الوظائف منها صنع الدهون والكاربوهيدرات وطيّ البروتينات في أكياس تسمى صهاريج ونقل هذه البروتينات المخلّقة إلى جهاز غولجي، وتسبب الاضطرابات في عملها استجابة إجهاد الشبكة الإندوبلازمية وهي حالة يكون فيها تطوي البروتينات بطيء مما يؤدي إلى زيادة البروتينات غير المتطوية، حيث يعتبر هذا الضغط سببا محتملا في أمراض نقص التأكسج/نقص التروية، مقاومة الأنسولين واضطرابات أخرى.
تاريخ
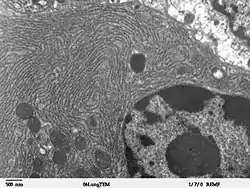
لوحِظت الشبكة الإندوبلازمية بواسطة المجهر الضوئي سنة 1897 من قبل غارنييه الذي صاغ مصطلح "الهيولى العاملة".[5][6] وتمت مشاهدة الأغشية الشريطة للشبكة بواسطة المجهر الإلكتروني لأول مرة سنة 1945 بواسطة كيث ر. بورتر، ألبير كلود، برودي ماسكرس وإيرنست ف. فولام.[7] ولا حقا، أُضيفت الكلمة "reticulum" والتي تعني "شبكة" بواسطة بواسطة بورتر سنة 1953 لوصف تركيب هذه الأغشية.[8]
البنية
البنية العامة للشبكة الإندوبلازمية عبارة عن شبكة من أغشية تسمى صهاريج، هذه البُنى الشبيهة بالأكياس مشدودة معا بواسطة الهيكل الخلوي. يحيط الغشاء اللبيدي الفوسفوري بالفراغ الصهريجي (أو اللمعة) وهو امتداد للفراغ النووي لكنه مفصول عن العصارة الخلوية. يمكن تلخيص وظيفة الشبكة الإندوبلازمية في تخليق وتصدير البروتينات والليبيدات الغشائية، لكنها تختلف حسب نوع الخلية ووظيفتها. يمكن أن تتغير كمية كل من الشبكة الإندوبلازمية الخشنة والملساء في الخلية ببطء من نوع لآخر، وذلك حسب تغيرات النشاطات الأيضية في الخلية. يمكن أن تشمل التحولات إدماج بروتينات جديدة في الغشاء بالإضافة إلى تغييرات بنيوية، قد يحدث تغيير في محتوى البروتين من دون تغيير بنيوي جليٍّ.[9][10]
الشبكة الإندوبلازمية الخشنة
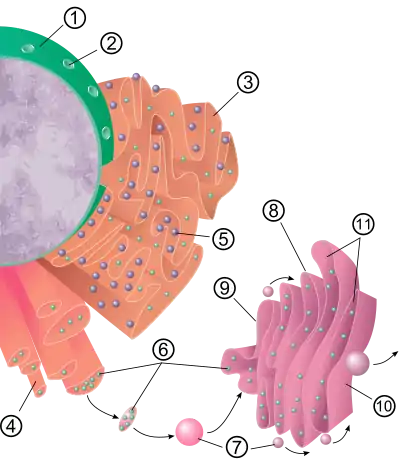
سطح الشبكة الإندوبلازمية الخشنة (وتسمى كذلك الشكبة الإندوبلازمية الحُبيبية) مرصّع بالريبوسومات المخلِّقة للبروتين وهذا يعطيها منظرا "خشنا" ومن هنا جاء اسمها.[11] موقع ارتباط الريبوسوم في الشبكة الإندوبلازمية هو المزفي،[12] إلا أن الريبوسومات ليست أجزاء مستقرة من بنية هذه العضية لأنها في حالة ارتباط وانفصال مستمرة بالغشاء. لا يرتبط الريبوسوم بالشبكة الخشنة سوى عند تشكل مركب بروتين-حمض نووي خاص في العصارة الخلوية، هذا المركب الخاص يتشكل حين يبدأ ريبوسومٌ حرٌ في ترجمة رنا رسول لبروتين هدفه المسار الإفرازي.[13] الأحماض الأمينية 5-30 الأولى تشفر ببتيد إشعاري وهو رسالة جزيئية يتم التعرف عليها والارتباط بها من قبل جسيم متعرف على الإشارة. تتوقف الترجمة ويرتبط مركب الريبوسوم بمزفي الشبكة الخشنة أين تتواصل الترجمة لتخليق بروتين وليد (جديد) على غشاء الشبكة أو داخل لمعتها. بعدها تتم معالجة البروتين داخل اللمعة بواسطة إنزيم (ببتيداز إشارة) الذي يزيل الببتيد الإشعاري. بعد هذه المرحلة يمكن أن تنفصل الريبوسومات وتعود للعصارة الخلوية، إلا أن الريبوسومات غير المترجِمة تُعرف ببقائها متصلة بالمزفيات.[14]
يشكل غشاء الشبكة الخشنة ملاءات كبيرة مزدوجة الأغشية تقع قرب الطبقة الخارجية للغلاف النووي وتمتد منها.[15] هذه الملاءات مزدوجة الغشاء مُحزّمة ومتصلة ببعضها بواسطة منحدرات لولبية يمينة أو يسارية، تسمى منحدرات تيراساكي، وهذا يؤدي إلى تشكل بنية تشبه موقف سيارات متعدد الطوابق.[16][17] رغم أنه لا يوجد غشاء مستمر بين الشبكة الإندوبلازمية وجهاز غولجي، إلا أن حويصلات غشائية تقوم بنقل البروتينات بين هذين الحيزين الخلويين.[18] الحويصلات محاطة ببروتينات تكسية تسمى COPI وCOPII، يستهدف COPII الحويصلات الذاهبة لجهاز غولجي ويسم COPI هذه الحويصلات لتتم إعادتها إلى الشبكة. تعمل الشبكة الإندوبلازمية في توافق مع جهاز غولجي لتوجيه البروتينات الجديدة إلى وجهاتها الصحيحة. توجد طريقة ثانية للنقل من الشبكة الإندوبلازمية تتم بواسطة مناطق تسمى مواقع اتصال غشائية، يكون فيها غشائي الشبكة الإندوبلازمية وعضيات أخرى قريبين جدا من بعضهما وهذا يسمح بنقل الليبيدات وجزيئات صغيرة أخرى.[19][20]
تقوم الشبكة الإندوبلازمية الخشنة بالعديد من الأدوار المفتاحية:
- تخليق الإنزيمات اليحلولية بإضافة علامة مانوز 6-فوسفات إلى الوجه مقرون لجهاز غولجي.[بحاجة لمصدر]
- تخليق البروتينات المُفرَزَة، سواء المفرَزة جوهريا من دون واسم أو المفرَزة بطريقة منتظمة بتوسط الكلاثرين وأحماض أمينية مقترنة في الببتيد الإشعاري.
- البروتينات الغشائية المدمجة التي تبقى مدمجة في الغشاء حين تغادر الحويصلات وترتبط بأغشية جديدة. بروتينات راب أساسية في استهداف الغشاء، بروتينات SNAP 25 وSNARE أساسية في مرحلة الاندماج
- الغلكزة الأولية بينما يستمر تخليق وتجميع البروتين، وتتم بالارتباط بذرة النيتروجين (في حين تتم الغلكزة المرتبطة بالأكسجين في جهاز غولجي).
- الغلكزة المرتبطة بالنيتروجين: إن كان البروتين متطويا بشكل صحيح، تتعرف ناقلة قليل الساكاريل على تسلسل الأحماض الأمينية NXS أوNXT (مع كون الوحدات S/T مفسفرة) وتضيف عمودا فقريا من 14 جزيئة سكر ( جزيئتي N-أسيتيل غلوكوز أمين، 9 جزيئات مانوز متفرع وثلاث جزيئات جلوكوز في النهاية) للسلسة الجانبية النيتروجينية للأسباراجين.
الشبكة الإندوبلازمية الملساء
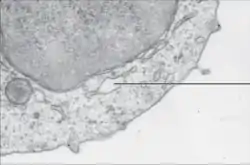
الشبكة الإندوبلازمية الملساء قليلة في معظم الخلايا، وبدل ذلك توجد مناطق في الشبكة ملساء جزئيا وخشنة جزئيا وتسمى هذه المناطق بالشبكة الإندوبلازمية الانتقالية وذلك لاحتوائها على مواقع لمغادرة الشبكة الإندوبلازمية، في هذه المناطق تنفصل الحويصلات الناقلة التي تحتوي على ليبيدات وبروتينات مخلَّقة عن الشبكة وتبدأ في التحرك نحو جهاز غولجي. يمكن أن يكون لدى الخلايا المتخصصة الكثير من الشبكة الملساء وفي هذه الخلايا يكون للشبكة الملساء وظائف متعددة،[9] فهي تخلِّق الليبيدات والليبيدات الفوسفورية والستيرويدات. الخلايا التي تُفرز هذه النواتج مثل خلايا الخصية والمبيض والغدة الدهنية لديها شبكة إندوبلازمية ملساء وفيرة.[21] تقوم الشبكة الملساء أيضا بأيض الكربوهيدرات، إزالة سمية النواتج الطبيعية للأيض والكحول والأدوية، وتقوم بوصل المستقبلات على بروتينات الخلية الغشائية وبأيض الستيرويدات.[22] وتقوم في الخلايا العضلية بتنظيم تركيز أيونات الكالسيوم. تتواجد الشبكة الملساء في أنواع مختلفة من الخلايا (الحيوانية والنباتية)، وتقوم بوظائف مختلفة في كل واحدة منها. تحتوي الشبكة الملساء على إنزيم غلوكوز 6-فسفاتاز الذي يحول غلوكوز 6-فوسفات إلى غلوكوز، وهي مرحلة في عملية استحداث الجلوكوز. الشبكة الملساء مرتبطة بالغشاء النووي وتتكون من أنيببات تتموضع بالقرب من محيط الخلية، تتفرع هذه الأنيببات أحيانا مشكلة تفرعات شبكية المظهر.[15] في بعض الخلايا، توجد مناطق متوسعة تشبه أكياس الشبكة الإندوبلازمية الخشنة. تفرعات الشبكة الإندوبلازمية الملساء تسمح بزيادة المساحة المخصصة لتخزين الإنزيمات الضرورية ونواتج هذه الإنزيمات.
الشبكة الإندوبلازمية العضلية
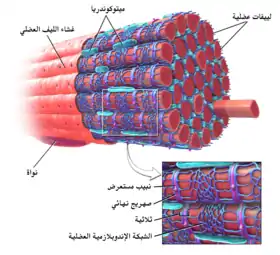
الشبكة الإندوبلازمية العضلية: مشتقة من الكلمة الإريقية σάρξ بمعنى "لحم"، وهي شبكة إندوبلازمية ملساء تتواجد في الخلايا العضلية. الفرق البنيوي الوحيد بين هذه العضية والشبكة الإندوبلازمية هو خليط وأنواع البروتينات التي تحتوي عليها، والتي تكون مدمجة في غشاء كلا النوعين وتتحرك داخل حدود اللمعات الخاصة بها. هذا الفرق الأساسي مؤشر على الوظائف التي تمارسها كل شبكة منهما: فالشبكة الإندوبلازمية تخلِّق الجزيئات، في حين أن الشبكة الإندوبلازمية العضلية تخزن أيونات الكالسيوم وتضخها خارجا إلى الهيولى العضلية حين يتحفز الليف العضلي.[23][24] بعد تحررها من الشبكة الإندوبلازمية العضلية، تتآثر أيونات الكالسيوم مع بروتينات انقباضية تستخدم الـATP لتقصير الليف العضلي. تلعب الشبكة الإندوبلازمية العضلية دورا كبيرا في انقباض العضلات.[25]
الوظائف
تقوم الشبكة الإندوبلازمية بالعديد من الوظائف، بما في ذلك طيّ البروتينات في أكياس تسمى صهاريج ونقل هذه البروتينات المخلّقة إلى جهاز غولجي. يحدث التطوي الصحيح للبروتينات بفضل عدة شابرونات بروتينية خاصة بالشبكة الإندوبلازمية منها: مخلقة ثنائي كبريتيد البروتين (PDI) وERp29 وعضو عائلة Hsp70 البروتين BiP/Grp78 والكالنيكسين والكالريتكولين وعائلة مصاوغة بيبتيديل بروبيل. البروتينات المتطوية وحدها من ينتقل من الشبكة إلى جهاز غولجي، في حين تُسبِّب البروتينات غير المتطوية استجابة البروتين غير المتطوي وهي استجابة للإجهاد في الشبكة الإندوبلازمية. يمكن أن تقود اضطرابات في تنظيم الخزدلة، تنظيم الكالسيوم، حرمان الغلوكوز، والعدوى الفيروسية[26] أو التعبير الزائد عن البروتينات[27] إلى استجابة إجهاد الشبكة الإندوبلازمية وهي حالة يكون فيها تطوي البروتينات بطيء مما يؤدي إلى زيادة البروتينات غير المتطوية. يتجلى هذا الضغط كسبب محتمل في تضرر نقص التأكسج/نقص التروية، مقاومة الأنسولين واضطرابات أخرى.[28]
نقل البروتين
تُنقَل البروتينات المفرزة -في الغالب بروتينات سكرية- على طول غشاء الشبكة الإندوبلازمية، وتوسم البروتينات التي تُنقل بواسطة الشبكة الإندوبلازمية عبر الخلية بواسِم عنوانٍ يسمى تسلسل إشعاري. تحتوي النهاية الأمينية لسلسة عديد الببتيد على عدة أحماض أمينية تعمل كواسم عنوان والذي يُزال حين يصل عديد الببتيد إلى وجهته. تصل الببتيدات الوليدة إلى الشبكة الإندوبلازمية عبر المزفيات وهي مركب عديد بروتين مدمج في الغشاء. تُحزم البروتينات التي وجهتها خارج الشبكة الإندوبلازمية في حويصلات ناقلة وتتحرك على طول الهيكل الخلوي إلى غايتها. في الأرومة الليفية البشرية، دائما ما تتوزع الشبكة مع الأنيببات الدقيقة وفك بلمرة هذه الأنيببات يسبب تكدسها مع المتقدرات التي هي بدورها متصلة بالشبكة الإندوبلازمية.[29]
الشبكة الإندوبلازمية جزء من مسار تصنيف البروتين، وهي نظام النقل في خلايا حقيقيات النوى. تُستبقى معظم بروتينات الشبكة الإندوبلازمية المقيمة داخلها من خلال نمط استبقاء، هذا النمط عبارة عن أربع أحماض أمينية في نهاية تسلسل البروتين. أكثر تسلسلات الاستبقاء شيوعا هما KDEL بالنسبة للبروتينات التي تُستبقى في اللمعة وKKXX بالنسبة للبروتينات غبر الغشائية.[30] إلا أنه تحدث تغيرات لـKDEL وKKXX ويمكن أن تقوم تسلسلات أخرى باستبقاء البروتينات في الشبكة. لا يُعرف إن كان من الممكن أن تقود هذه التغيرات إلى توضعات فرعية في الشبكة. توجد ثلاث مستقبلات من KDEL (2 ،1 و3) في خلايا الثدييات ولديها درجة عالية من التعرف على التسلسل، والاختلافات الوظيفية بين هذه المستقبلات لم يتم تحديدها بعد.[31]
الأهمية السريرية
يقود الشذوذ في بروتين XBP1 إلى استجابة إجهاد شبكة إندوبلازمية متزايدة وهذه الاستجابة تسبِّبُ قابليةً أعلى لعمليات الالتهاب التي يمكن أن تساهم حتى في مرض الألزهايمر،[32] وفي القولون تم ربط شذوذ XBP1 مع أمراض التهاب الأمعاء بما في ذلك داء كرون.[33]
استجابة البروتين غير المتطوي (UPR) هي استجابةُ إجهادٍ خلويةٍ متعلقة بالشبكة الإندوبلازمية.[34] تفعَّل هذه الاستجابة كرد على تراكم البروتينات غير المتطوية والمتطوية بشكل خاطئ في لمعة الشبكة الإندوبلازمية، ووظيفتها هي استعادة الوظيفة الطبيعية للخلية عبر وقف ترجمة البروتينات، تحليل وفك البروتينات المتطوية بشكل خاطئ، وتفعيل مسارات التأشير التي تؤدي إلى زيادة إنتاج الشابرونات المساهِمة في تطوي البروتين. تم اعتبار فرط النشاط المستمر لاستجابة البروتين غير المتطوي من أسباب أمراض البريون وكذلك العديد من أمراض التحلل العصبي الأخرى، ومنع استجابة البروتين غير المتطوي قد يصبح علاجا لهذه الأمراض.[35]
المراجع
- المعجم الطبي الموحد [وصلة مكسورة] نسخة محفوظة 16 أبريل 2016 على موقع واي باك مشين.
- الموسوعة الريفية
- كتاب الأحياء للصف الثاني ثانوي علمي؛ المملكة الأردنية الهاشمية؛ طبعة عام 1997، صفحة 11
- مقرر علم الأحياء، الصف الأول متوسط، صفحة 63، دولة العراق.
- Garnier, C. 1897. Les filaments basaux des cellules glandulaires. Note préliminaire. Bibliographie anatomique, 5, 278–289.
- Buvat, R. 1963. Electron microscopy of plant protoplasm. Int. Rev. Cytol. 14: 41–155. link نسخة محفوظة 2018-04-07 على موقع واي باك مشين..
- "A study of tissue culture cells by electron microscopy: methods and preliminary observations". The Journal of Experimental Medicine. 81 (3): 233–46. March 1945. doi:10.1084/jem.81.3.233. PMC 2135493. PMID 19871454. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Porter, K. R. Observations on a submicroscopic basophilic component of cytoplasm. J Exp Med. 1953 May; 97(5):727–750. link نسخة محفوظة 2015-11-21 على موقع واي باك مشين..
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2002). Molecular biology of the cell (الطبعة 4th). New York: Garland Science. ISBN 978-0-8153-3218-3. مؤرشف من الأصل في 03 أكتوبر 2017. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cooper GM (2000). The cell: a molecular approach (الطبعة 2nd). Washington (DC): ASM Press. ISBN 978-0-87893-106-4. مؤرشف من الأصل في 11 مارس 2014. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "reticulum". The Free Dictionary. مؤرشف من الأصل في 20 نوفمبر 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "A mammalian homolog of SEC61p and SECYp is associated with ribosomes and nascent polypeptides during translocation". Cell. 71 (3): 489–503. October 1992. doi:10.1016/0092-8674(92)90517-G. PMID 1423609. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lodish H, et al. (2003). Molecular Cell Biology (الطبعة 5th). W. H. Freeman. صفحات 659–666. ISBN 978-0-7167-4366-8. مؤرشف من الأصل في 26 يناير 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The fate of membrane-bound ribosomes following the termination of protein synthesis". The Journal of Biological Chemistry. 275 (43): 33820–7. October 2000. doi:10.1074/jbc.M004462200. PMID 10931837. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Rough sheets and smooth tubules". Cell. 126 (3): 435–9. August 2006. doi:10.1016/j.cell.2006.07.019. PMID 16901774. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Stacked endoplasmic reticulum sheets are connected by helicoidal membrane motifs". Cell. 154 (2): 285–96. July 2013. doi:10.1016/j.cell.2013.06.031. PMC 3767119. PMID 23870120. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Terasaki spiral ramps in the rough endoplasmic reticulum". Physical Review Letters. 113 (18): 188101. October 2014. doi:10.1103/PhysRevLett.113.188101. PMID 25396396. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Endoplasmic reticulum. (n.d.). McGraw-Hill Encyclopedia of Science and Technology. Retrieved September 13, 2006, from Answers.com Web site: "Answers - the Most Trusted Place for Answering Life's Questions". مؤرشف من الأصل في 16 نوفمبر 2006. اطلع عليه بتاريخ 13 سبتمبر 2006. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Short-range intracellular trafficking of small molecules across endoplasmic reticulum junctions". Trends in Cell Biology. 14 (9): 483–90. September 2004. doi:10.1016/j.tcb.2004.07.017. PMID 15350976. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Inter-organelle membrane contact sites: through a glass, darkly". Current Opinion in Cell Biology. 18 (4): 371–8. August 2006. doi:10.1016/j.ceb.2006.06.011. PMID 16806880. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Functions of Smooth ER". University of Minnesota Duluth. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Intracellular cholesterol transport". The Journal of Clinical Investigation. 110 (7): 891–8. October 2002. doi:10.1172/JCI16500. PMC 151159. PMID 12370264. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Crystal structure of the calcium pump of sarcoplasmic reticulum at 2.6 A resolution". Nature. 405 (6787): 647–55. June 2000. doi:10.1038/35015017. PMID 10864315. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Goodman SR (2007-11-26). Medical Cell Biology (الطبعة 3rd). Academic Press. صفحة 69. ISBN 9780080919317. مؤرشف من الأصل في 11 مايو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Martini F, Nath J, Bartholomew E (2014). Fundamentals of Anatomy and Physiology (الطبعة 10th). ISBN 978-0321909077. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Endoplasmic reticulum stress: cell life and death decisions". The Journal of Clinical Investigation. 115 (10): 2656–64. October 2005. doi:10.1172/JCI26373. PMC 1236697. PMID 16200199. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Development of a novel ER stress based selection system for the isolation of highly productive clones". Biotechnology and Bioengineering. 109 (10): 2599–611. October 2012. doi:10.1002/bit.24527. PMID 22510960. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Endoplasmic reticulum stress links obesity, insulin action, and type 2 diabetes". Science. 306 (5695): 457–61. October 2004. doi:10.1126/science.1103160. PMID 15486293. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Interrelationships of endoplasmic reticulum, mitochondria, intermediate filaments, and microtubules--a quadruple fluorescence labeling study". Biochemistry and Cell Biology. 70 (10–11): 1174–86. 1992. doi:10.1139/o92-163. PMID 1363623. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "KDEL and KKXX retrieval signals appended to the same reporter protein determine different trafficking between endoplasmic reticulum, intermediate compartment, and Golgi complex". Molecular Biology of the Cell. 14 (3): 889–902. March 2003. doi:10.1091/mbc.E02-08-0468. PMC 151567. PMID 12631711. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "A molecular specificity code for the three mammalian KDEL receptors". The Journal of Cell Biology. 179 (6): 1193–204. December 2007. doi:10.1083/jcb.200705180. PMC 2140024. PMID 18086916. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The ER stress factor XBP1s prevents amyloid-beta neurotoxicity". Human Molecular Genetics. 20 (11): 2144–60. June 2011. doi:10.1093/hmg/ddr100. PMC 3090193. PMID 21389082. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "XBP1 links ER stress to intestinal inflammation and confers genetic risk for human inflammatory bowel disease". Cell. 134 (5): 743–56. September 2008. doi:10.1016/j.cell.2008.07.021. PMC 2586148. PMID 18775308. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Walter, Peter. "Peter Walter's Short Talk: Unfolding the UPR". iBiology. مؤرشف من الأصل في 16 نوفمبر 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Oral treatment targeting the unfolded protein response prevents neurodegeneration and clinical disease in prion-infected mice". Science Translational Medicine. 5 (206): 206ra138. October 2013. doi:10.1126/scitranslmed.3006767. PMID 24107777. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة علم الأحياء
بوابة علم الأحياء بوابة طب
بوابة طب- بوابة علم الأحياء الخلوي والجزيئي