مريكز
في علم الأحياء الخلوي؛ المريكز هو عضية خلوية أسطوانية الشكل تظهر على شكل ازواج،[1] تتكون أساسا من بروتين يسمى تيوبيولين. توجد المريكزات في معظم الخلايا حقيقيات النوى. في العادة يتواجد زوج من المريكزات داخل كل خلية وتحيط بهما كتلة عديمة الشكل من مادة كثيفة تشكل هيكل ما يُسمى بالجسيم المركزي.
المريكزات موجودة في خلايا معظم حقيقيات النوى فعلى سبيل المثال هي موجودة في الخلايا الحيوانية، ومع ذلك فهي غائبة عن الصنوبريات (المخروطيات على سبيل المثال لا الحصر) كما لا تتواجد في بعض النباتات المزهرة (كاسيات البذور مثلا) ومعظم الفطريات. تتواجد المريكزات فقط في الأمشاج الذكرية لكل من النباتات الكاريانية، البريويات، النباتات الوعائية بدون بذور.[2][3]
عادة ما يتكون المريكز من تسع مجموعات من الأنيبيب الدقيق على شاكلة ثلاثة توائم مرتبة في أسطوانة. الانحرافات في شكل المريكز وهيكله نادرة لكنها قد تحدث في بعض الحالات خاصة عندما يتعلق الأمر بحيوان السرطانات، ذبابة الفاكهة سوداء البطن والربداء الرشيقة.[4][5]

تتمثل المهمة الرئيسية للمريكزات في إنتاج ما يُعرف بالأهداب وذلك خلال الطور البيني؛ كما تقوم بأدوار ثانوية أخرى من بينها المساعدة على تشكيل المغزل أثناء الانقسام الخلوي.
التاريخ
يُعد العالِم إدوارد فان بيندين أول من دوَّن ملاحظات حول المريكزات عام 1883.[6] في عام 1895؛ تعمَّق تيودور بوفيري في الموضوع فسمَّى ما رآه بـ "المريكز".[7][8] في عام 1950 عمل العالِمان إتيان دي هارفن وجوزيف على محاولة التعرَّف على المريكز ووضع ورقية بحثية شاملة حول هذا العُضي؛ حينها تمكنا من استخلاص العديد من المعلومات من بينها.[9][10]
دوره في انقسام الخلية

تشارك المريكزات في تنظيم الانقسامات الاختزالية كما تُساعد في الانتهاء من الانقسام السيتوبلازمي.[11] كان يُعتقد في السابق أن هناك حاجة ماسَّة للمريكزات حتى تستطيع الخلايا الحيوانية التكاثر من خلال الانقسام الاختزالي، بيد أن أكثر التجارب الأخيرة أثبتت أن المريكزات في الخلايا التي تم إزالتها واجتثاها عن طريق الليزر لم تتأثر بل واصلت الخلية تطورها و"زحفها" نحو مرحلة الطور البيني دون الحاجة لتلك المريكزات.[12] بالإضافة إلى ذلك، فالذباب المتحول لا يتوفر على مريكزات وبالرغم من ذلك فهو يتطور بشكل طبيعي، لكن وفي المقابل فخلايا كبار الذباب لا تتوفر على أسواط أو أهداب ونتيجة لذلك فإنها تموت بعد فترة قصيرة من الولادة.[13] هذا وتجدر الإشارة إلى أن المريكزات يُمكنها تكرار نفسها بشكل ذاتي خلال الانقسام الخلوي.
التنظيم الخلوي
المريكزات هي جزء مهم جدا من الجسيم المركزي الذي يُشارك في تنظيم الأنيبيب الدقيق في السيتوبلازم.[14][15] وتظهر أهمية المريكزات في أن موقفها يُحدد موقف النواة وبالتالي فالمريكزات تلعب دورا حاسما في الترتيب المكاني للخلية.

الخصوبة
تلعب المريكزات دورا هاما من خلال شغل وظيفتين تتعلقان بشكل مباشر وغير مباشر بالحيوانات المنوية:[16]
- تُؤثر المريكزات على شكل الحيوانات المنوية كما تُؤثر على السوط (الجهاز الحركي للخلية) وبالتالي فهي تُؤثر على حركة الحيوانات المنوية.
- تطور الجنين بعد الإخصاب.
دور آخر
في الكائنات الحية التي تتوفر على أسواط وأهداب، يتم تحديد مكان هذان العضيتان من المريكز الأم الذي يصبح فيما بعد جسم قاعدي. إن عدم قدرة الخلايا على استخدام المريكزات لتقوم الأهداب والأسواط بوظيفتها يزيد من احتمالية الإصابة بأمراض قد تكون وراثية، ولا سيما عندما تكون المريكزات عاجزة. في الآونة الأخيرة تم ربط متلازمة ميكل-غروبر بعجز هذه المريكزات.[17]
المريكز عند الثدييات
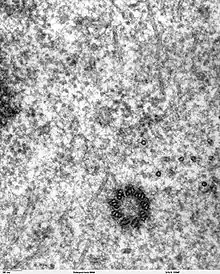
التوجه السليم للأهداب عبر المريكز باتجاه المواقع الخلفية الجنينية أو باتجاه الخلايا أمر معقد وبالغ الأهمية باعتبار أنه يُنشئ التباين بين كل من اليسار واليمين عند الثدييات.[18]
ازدواجية المريكز
قبل تضاعف الحمض النووي الريبوزي منقوص الأكسجين؛ تحتوي الخلايا على مُريكزان، الأكبر يُطلق عليه اسم المريكز الأم أما الثاني _الأصغر_ فيُطلق عليه اسم المريكز الابن. أثناء دورة انقسام الخلية ينمو مريكز جديد في نهاية قريبة من كل من المريكز الأم والمريكز الابن؛ وبالتالي يشتكل زوجين من كل مريكز (عندما يتم تجميعهم نحصل على زوجين أم وزوجين ابن). بصفة عامة تكون المريكزات متعامدة وتبقى على هذه الوضعية إلى حين الانقسام المتساو. عند هذه النقطة ينفصل المريكز الأم عن المريكز الابن بالاعتماد على إنزيم يسمى سيباراز (بالإنجليزية: separase)[lower-alpha 1].[19]
أما المريكزين في الجُسَيِّم المركزي فيمكثا مرتبطين، حيث أن المريكز الأم يأخد دور القاصي في حين يأخد المريكز الابن دور الداني. هذا وتجدر الإشارة إلى أن كل خلية بنت تشكلت بعد الانقسام الخلوي ترث واحدة من هذه الأزواج، فتبدأ المريكزات بتكرار نفسها حال تطابق الحمض النووي.
الأصل
آخر سلف مشترك لجميع حقيقيات النوى كان مهدبا ويحتوي على مريكزات خلوية، لكن وفي المقابل فهناك بعض الأنساب من حقيقيات النوى على غرار النباتات الجنينية والتي لا تحتوي على مريكزات إلا في الأمشاج الذكرية. ليس هذا فقط فلا وجود للمريكزات في كل من خلايا المخروطيات والنباتات المزهرة التي لا تملك الأهداب.[20] وبالتالي فمن غير الواضح ما إذا كان أول آخر سلف مشترك لحقيقيات النوى يحوي مريكزات أم لا.[21][22] لكن وبصفة عامة فهناك بعض الجينات المطلوبة لنمو المريكزات؛ وبعضها _أي بعض الجينات_ يوجد فقط في حقيقيات النوى ولا يوجد في البكتيريا أو البكتيريا القديمة.
انظر أيضًا
- خلية حيوية
ملاحظات
- تم تسمية الاسم بهذا الاسم الذي يُترجم حرفيا لـ الفاصل باعتبار دوره، حيث يقوم بالفصل بين المريكزين
المراجع
- Eddé, B; Rossier, J; Le Caer, JP; Desbruyères, E; Gros, F; Denoulet, P (1990). "Posttranslational glutamylation of alpha-tubulin". Science. 247 (4938): 83–5. Bibcode:1990Sci...247...83E. doi:10.1126/science.1967194. PMID 1967194. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Quarmby, LM; Parker, JD (2005). "Cilia and the cell cycle?". The Journal of Cell Biology. 169 (5): 707–10. doi:10.1083/jcb.200503053. PMID 15928206. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Silflow, CD; Lefebvre, PA (2001). "Assembly and motility of eukaryotic cilia and flagella. Lessons from Chlamydomonas reinhardtii". Plant Physiology. 127: 1500–1507. doi:10.1104/pp.010807. PMID 11743094. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Delattre, M; Gönczy, P (2004). "The arithmetic of centrosome biogenesis" (PDF). Journal of Cell Science. 117 (Pt 9): 1619–30. doi:10.1242/jcs.01128. PMID 15075224. مؤرشف من الأصل (PDF) في 18 أغسطس 2017. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Leidel, S.; Delattre, M.; Cerutti, L.; Baumer, K.; Gönczy, P (2005). "SAS-6 defines a protein family required for centrosome duplication in C. elegans and in human cells". Nature Cell Biology. 7 (2): 115–25. doi:10.1038/ncb1220. PMID 15665853. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wunderlich, V. (2002). "JMM - Past and Present". Journal of Molecular Medicine. 80 (9): 545–548. doi:10.1007/s00109-002-0374-y. PMID 12226736. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Boveri, T. Ueber das Verhalten der Centrosomen bei der Befruchtung des Seeigel-Eies nebst allgemeinen Bemerkungen über Centrosomen und Verwandtes. Verh. d. Phys.-Med. Ges. zu Würzburg, N. F., Bd. XXIX, 1895. link.
- Boveri, T. (1901). Zellen-Studien: Uber die Natur der Centrosomen. IV. Fischer, Jena. link.
- Wolfe, Stephen L. (1977). Biology: the foundations (الطبعة First). Wadsworth. مؤرشف من الأصل في 15 ديسمبر 2016. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Vorobjev, I. A.; Nadezhdina, E. S. (1987). "The Centrosome and Its Role in the Organization of Microtubules". International Review of Cytology. 106: 227–293. doi:10.1016/S0074-7696(08)61714-3. ISBN 978-0-12-364506-7. PMID 3294718. الوسيط
|CitationClass=تم تجاهله (مساعدة). See also de Harven's own recollections of this work: de Harven, Etienne (1994). "Early observations of centrioles and mitotic spindle fibers by transmission electron microscopy". Biol Cell. 80 (2–3): 107–109. doi:10.1111/j.1768-322X.1994.tb00916.x. PMID 8087058. مؤرشف من الأصل في 13 ديسمبر 2019. الوسيط|CitationClass=تم تجاهله (مساعدة) - Salisbury, JL; Suino, KM; Busby, R; Springett, M (2002). "Centrin-2 is required for centriole duplication in mammalian cells". Current Biology. 12 (15): 1287–92. doi:10.1016/S0960-9822(02)01019-9. PMID 12176356. الوسيط
|CitationClass=تم تجاهله (مساعدة) - La Terra, S; English, CN; Hergert, P; McEwen, BF; Sluder, G; Khodjakov, A (2005). "The de novo centriole assembly pathway in HeLa cells: cell cycle progression and centriole assembly/maturation". The Journal of Cell Biology. 168 (5): 713–22. doi:10.1083/jcb.200411126. PMID 15738265. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Basto, R; Lau, J; Vinogradova, T; Gardiol, A; Woods, CG; Khodjakov, A; Raff, JW (2006). "Flies without centrioles". Cell. 125 (7): 1375–86. doi:10.1016/j.cell.2006.05.025. PMID 16814722. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Feldman, JL; Geimer, S; Marshall, WF (2007). "The mother centriole plays an instructive role in defining cell geometry". PLoS Biology. 5 (6): e149. doi:10.1371/journal.pbio.0050149. PMID 17518519. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Beisson, J; Wright, M (2003). "Basal body/centriole assembly and continuity". Current Opinion in Cell Biology. 15 (1): 96–104. doi:10.1016/S0955-0674(02)00017-0. PMID 12517710. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Avidor-Reiss, T., Khire, A., Fishman, E. L., & Jo, K. H. (2015). Atypical centrioles during sexual reproduction. Frontiers in cell and developmental biology, 3, 21. Chicago
- Cui, Cheng; Chatterjee, Bishwanath; Francis, Deanne; Yu, Qing; SanAgustin, Jovenal T.; Francis, Richard; Tansey, Terry; Henry, Charisse; Wang, Baolin (2011). "Disruption of Mks1 localization to the mother centriole causes cilia defects and developmental malformations in Meckel-Gruber syndrome". Dis Model Mech. 4 (1): 43–56. doi:10.1242/dmm.006262. PMID 21045211. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Babu, Deepak; Roy, Sudipto (2013-05-01). "Left–right asymmetry: cilia stir up new surprises in the node". Open Biology (باللغة الإنجليزية). 3 (5): 130052. doi:10.1098/rsob.130052. ISSN 2046-2441. PMID 23720541. مؤرشف من الأصل في 11 أغسطس 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Tsou, MF; Stearns, T (2006). "Mechanism limiting centrosome duplication to once per cell cycle". Nature. 442 (7105): 947–51. Bibcode:2006Natur.442..947T. doi:10.1038/nature04985. PMID 16862117. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Marshall, W.F. (2009). "Centriole Evolution". Current Opinion in Cell Biology. 21 (1): 14–19. doi:10.1016/j.ceb.2009.01.008. PMID 19196504. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bornens, M.; Azimzadeh, J. (2007). "Origin and Evolution of the Centrosome". Eukaryotic Membranes and Cytoskeleton. 607. صفحات 119–129. doi:10.1007/978-0-387-74021-8_10. ISBN 978-0-387-74020-1. PMID 17977464. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Rogozin, I. B.; Basu, M. K.; Csürös, M.; Koonin, E. V. (2009). "Analysis of Rare Genomic Changes Does Not Support the Unikont-Bikont Phylogeny and Suggests Cyanobacterial Symbiosis as the Point of Primary Radiation of Eukaryotes". Genome Biology and Evolution. 1: 99–113. doi:10.1093/gbe/evp011. PMID 20333181. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علم الأحياء
بوابة علم الأحياء
