انقباض عضلي
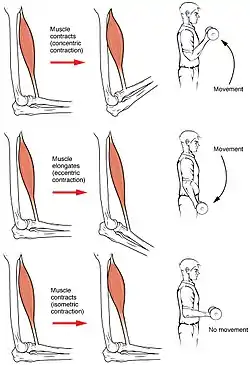
الانقباض العضلي هو الحركة المعتادة للجسم وهي تعتمد على انقباض ثم تراخي للعضلات. ينشأ انقباض العضلة من انزياح جزيئات الميوسين على فتيلات جزيئات الأكتين في الخلية العضلية. جزيئات الميوسين والأكتين في الخلية العضلية تتداخل في بعضها البعض كأسنان المشط، وعندما يحدث الشد بينهما وتقصر المسافة فهذا هو الانقباض ويتحرك العضو (مثل ثني الذراع أو حركة الرجل إلى الأمام) . وهكذا تنشأ حركات الجسم بواسطة عضلات مختلفة موزعة على أطرافه واعضائه. في الانقباض والتراخي تتعرض العضلة إلى القصر أو الاستطالة أو تبقى كما هي. هذا غير ما يسمى التقلص أو التشنج spasm، فالتقلص هو حالة اعياء للعضلة وأوجاع بسبب الإجهاد الزائد عن اللزوم، ويحتاج "لفكه" وقتا طويلا ربما أسبوع أو أكثر.
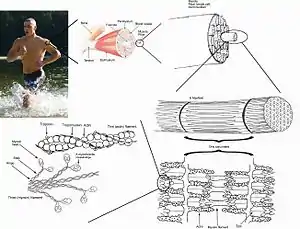
ويتم تحريك العضلات من خلال النظام العصبي المركزي. ويقوم المخ بإرسال الإشارات، في شكل جهود الفعل، من خلال النظام العصبي إلى الخلايا العصبية الحركية المستخدمة للتوصيل العصبي للعديد من الألياف العضلية (جزيئات الميوسين والأكتين، أنظر الشكل ) .
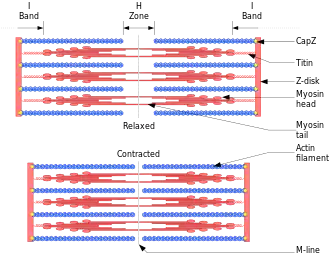
وصف آلية الانقباض
 مقالة مفصلة: نظرية الخيط المنزلق
مقالة مفصلة: نظرية الخيط المنزلق
نظرية انزلاق الفتيلات
طبقا لنظرية الانزلاق الفتيلي التي وضعها " أندرو هيكسلي" و "هيو هيكسلي" [1] عند الانقباض تنزلق فتيلات بروتين من دون تغير في طولها على بعضها البعض كأسنان المشط وتقصر بذلك طول العضلة. فتيلات البروتين عبارة عن جزيئات أكتين الفتيل الخارجي الرقيق وتنزلق عليها فتيلات داخلية سميكة من جزيئات ميوسين (بروتين) وبهذا تعمل على تقصير المسافة بينها، ويحدث انقباض العضلة. تنشأ هذه الحركة من تغير في الشكلية الكيميائية للميوسين، وبالتالي تغير في شكل جزييء الميوسين. الميوسين يتكون من رؤوس صغيرة يمكنها تغيير زاويتها بالنسبة للجزع الطويل. ويمكن لتلك الرؤوس للميوسين أن تتعلق بفتيلات الأكتين وتنزلق عليها بفعل حركة تشبه التجديف. وينشأ انقباض الميوسين من نبضة عصبية. كما ينفك الميوسين عن الأكتين بواسطة طاقة يقدمها جزيء اسمه أدينوسين ثلاثي الفوسفات ATP . فإذا لم تتوفر تلك الطاقة فإن الجزيئات لا تنفصل عن بعضها البعض.
بالتفصيل فإن انقباض عضلة يُفسر بحدوث ما يسمى "دورة قنطرة عرضية" (بمعني دورة شد واسترخاء ) بين فتيلات الميوسين والأكتين. ويأتي هذا الاسم من وظيفة رؤوس الميوسين وحركتها بين فتيلات الميوسين والأكتين.
- في حالة الهدوء (استرخاء العضلة) تكون فتيلة الأكتين في الخلية العضلية محزمة بفتيلات من التروبوميوسين وهي تغطي أماكن ارتباط رؤوس الميوسين على فتيل الأكتين.
- أدينوسين ثلاثي الفوسفات ATP - وهو مصدر الطاقة - يكون موجودا في داخل الميوسين، وتكون أوضاع الرؤوس بزاوية 90 درجة بالنسبة لجزع جزيء الميوسين.وتأتي نبضة عصبية تفرز أيونات الكالسيوم (Ca2+) في الميوسين. يتسبب هذا الإفراز للكاسيوم في فعلين: يُنشط الكالسيوم انزيمات رؤوس الميوسين التي تحوّل الأدينوسين ثلاثي الفوسفات ATP إلى ادينوسين ثنائي الفوسفات ADP وتحرر فوسفات Pi . تلك العملية تحتاج إلى أيونات مغنسيوم Mg2+ التي تقوم كعامل مساعد (مرافق) بفك رابطة الفوسفات من ATP. ومن جهة أخرى يترابط الكالسيوم على التروبونين الموجود على فتيل التروبوميوسين ويغير شكل التروبوميوسين بحيث يتعرى مكان الارتباط فيحدث تشابك بين الميوسين على الأكتين.
- بمجرد حدوث تشابك بين الميوسين والأكتين فإن جزيء الفوسفا ت الذي لا زال مرتبطا برأس الميوسين يعمل على الفور أيضا على تحرير أدينوسين ثنائي الفوسفات ADP . بذلك يتحول الجهد على الميوسين إلى حركة ميكانيكية. وتغير رؤوس الميوسين وضعها من زاوية 90 درجة إلى زاوية 45 درجة بالنسبة إلى جزع الميوسين ( هذه ضربة تشبه ضربات التجديف ) وتسمى تلك الحركة قوة دافعة، وتـُشد بذلك فتيلات الأكتين من اليمين واليسار إلى وسط حزمة الألياف العضلية Sarcomere.[2])
- تنتهي الدورة بذلك بحيث أن يتجمع ATP جديد على الميوسين. فتحل رأس الميوسين نفسها من فتيل
الأكتين ويعود هذين البروتينين إلى اتخاذ وضعيهما الأولي.
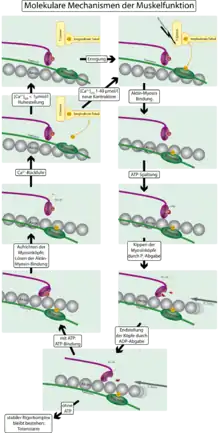
تستغرق تلك دورة القنطرة العرضية بين 10 إلى 100 ميكروثانية وهي تسحب الفتيلة مقدار 10 إلى 20 نانومتر ، وهذا يشكل تغيرا في طولها بمقدار 1%. ولكي يحدث تغير أكبر في الطول فإن الدورة تتكرر وتتبعها دورات عديدة. وبعد نحو 50 دورة شد واسترخاء ستنتج إزاحة كلية لحزمة الألياف العضلية بمقدر 50% من طولها أثناء الهدوء خلال جزء من الثانية.
هذا الوصف يخص ما يجري في خلية عضلية، والخلايا العضلية تكون متراصة طوليا بحيث تعمل في نفس الاتجاه لتقوم سويا بالحركة. فعندما تأتي إشارة من الدماغ إلى العضلة فإن الإشارة تتفرع وتتوزع بالأعصاب على جميع الخلايا العضلية، وبذلك تعمل مجموعة الخلايا العضلية جماعيا وتحدث حركة العضو، سواء يد أو ساعد أو رجل.
الفقاريات
في الحيوانات الفقارية، ثمّة ثلاثة أنواع من الأنسجة العضلية: الهيكلية والملساء والقلبية. تشكل الأنسجة العضلة الهيكلية غالبية كتلة العضلات في الجسم وهي مسؤولة عن النشاط الحركي. تشكل العضلات الملساء الأوعية الدموية والسبيل الهضمي وغيرها من المناطق في الجسم التي تقوم بانقباضات مستمرة. ألياف العضلة القلبية تشكل القلب الذي يضخ الدم. تسمى العضلات الهيكلية والعضلات القلبية عضلة مخططة بسبب مظهرها المخطط تحت المجهر، ويعود ذلك إلى النمط المتناوب المنتظم بشدة لحزم إيه وحزم آي.
العضلة الهيكلية
باستثناء ردود الفعل الانعكاسية، تحدث جميع انقباضات العضلات الهيكلية نتيجة لجهود واعية ناشئة في الدماغ. ويرسل الدماغ إشارات كهروكيميائية عبر الجهاز العصبي إلى العصبون الحركي الذي يعصب عدة ألياف عضلية.[3] وفي حالة بعض المنعكسات، يمكن أن تنشأ إشارة التقلص في النخاع الشوكي من خلال حلقة تلقيم راجع مع المادة الرمادية. أما النشاطات الأخرى مثل الحركة والتنفس والمضغ، فهي تنطوي على جانب انعكاسي: إذ يمكن أن يبدأ التقلص بوعي أو بدون وعي.
الوصل العصبي العضلي
الوصل العصبي لعضلي هو مشبك كيميائي يتكون من اتصال بين نهايات العصبون المحرك وألياف العضلات. وهو الموقع الذي ينقل فيه العصبون المحرك الإشارة إلى الليف عضلي ليبدأ تقلصه العضلي.[4] يبدأ تسلسل الأحداث التي تؤدي لإزالة استقطاب الألياف العضلية عند الوصل العصبي لعضلي بنشوء كمون العمل في جسم العصبون المحرك الذي ينتشر بعد ذلك بواسطة النقل القفزي على طول المحوار باتجاه الوصل العصبي العضلي. حالما يصل كمون العمل إلى الأزرار الانتهائية، يؤدي لتدفق شوارد الكالسيوم Ca2+ من الفالق المشبكي عبر قنوات الكالسيوم الفولطية إلى داخل الأزرار. يؤدي تدفق شوارد الكالسيوم Ca2+إلى اندماج الحويصلات المشبكية الحاوية على الناقل العصبي الكيميائي -الأستيل كولين- بجدار الأزرار الانتهائية محررة الأستيل كولين في الفالق المشبكي بين العصبون المحرك والوصل العصبي العضلي لألياف العضلة الهيكلية. ينتشر الأستيل كولين عبر المشبك العصبي ويرتبط بمستقبلات الأستيل الكولين النيكوتينية ويفعلها على الوصل العصبي العضلي. يؤدي تفعيل المستقبلات النيكوتينية لفتح قنوات الصوديوم/البوتاسيوم النوعية، ما يسبب تدفق الصوديوم إلى الداخل وتتدفق البوتاسيوم إلى الخارج. ونتيجة لذلك، ينعكس استقطاب الغشاء الخلوي، وسرعان ما تقفز الفولطية من كمون الراحة -90 مللي فولت إلى ما يساوي +75 مللي فولت عند دخول شوارد الصوديوم. ثم يدخل كمون الغشاء مرحلة فرط استقطاب عند خروج البوتاسيوم ثم يُعاد ضبطه مرة أخرى إلى كمون الراحة. هذا التقلب السريع يسمى كمون الصفيحة الانتهائية. تفتح القنوات الشاردية الفولطية للجدار الخلوي بجوار الصفيحة الانتهائية استجابة لكمون الصفيحة الانتهائية. هذه القنوات الفولطية هي قنوات نوعية للصوديوم والبوتاسيوم وتسمح فقط لعبور نوع واحد من الشوارد. هذه الموجة من حركة الشوارد تخلق كمون عمل ينتشر من الصفيحة الانتهائية المحركة في جميع الاتجاهات. إذا توقفت كمونات العمل عن الوصول، فإن الأستيل كولين يتوقف عن الخروج من الأزرار الانتهائية. أما الأستيل كولين المتبقي في الفالق المشبكي فيتفكك إما من خلال أنزيم أستيل كولين إستيراز فعال أو يُعاد امتصاصه بواسطة الزر المشبكي ولا يتبقى شيء منه وذلك ليحل محل الأستيل كولين المفكك.[5]
.jpg.webp)
انظر أيضًا
المراجع
- Hugh Huxley und Jean Hanson: Changes in the Cross-Striations of Muscle during Contraction and Stretch and their Structural Interpretation. In: طبيعة. Band 173, 1954, S. 973–976, doi:10.1038/173973a0
A. F. Huxley und R. Niedergerke: Structural Changes in Muscle During Contraction: Interference Microscopy of Living Muscle Fibres. In: Nature. Band 173, 1954, S. 971–973, doi:10.1038/173971a0.
vergl. dazu auch K. Maruyama (1995): Birth of the Sliding Filament Concept in Muscle Contraction. In: J Biochem. Band 117 (1): 1–6. - J. M. Berg, J. L. Tymoczko, L. Stryer: Biochemie. 6. Auflage. Spektrum Akademischer Verlag, Elsevier GmbH, München 2007; S. 1107f., ISBN 978-3-8274-1800-5.
- Tassinary; Cacioppo (2000). "The Skeletomotor system: surface electromyography". In Cacioppo, John T.; Tassinary, Luois G.; Berntson, Gary G. (المحررون). Handbook of Psychophysiology (الطبعة Second). Cambridge: Cambridge University Press. ISBN 978-0-521-62634-7. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Levitan, Irwin; Kaczmarek, Leonard (August 19, 2015). "Intercellular communication". The Neuron: Cell and Molecular Biology (الطبعة 4th). New York, NY: Oxford Univerty Press. صفحات 153–328. ISBN 978-0199773893. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Saladin, Kenneth S., Stephen J. Sullivan, and Christina A. Gan. Anatomy & Physiology: The Unity of Form and Function. 7th ed. New York: McGraw-Hill Education, 2015. Print.
 بوابة طب
بوابة طب بوابة علم وظائف الأعضاء
بوابة علم وظائف الأعضاء بوابة كمال الأجسام
بوابة كمال الأجسام
 صور وملفات صوتية من كومنز
صور وملفات صوتية من كومنز