بروتينات بولية رئيسية
البروتينات البولية الرئيسية MUPS ، والتي تعرف أيضًا باسم α2u-globulins، هي فصيلة بروتينات موجودة بكثرة في بول وإفرازات العديد من الحيوانات. توفر البروتينات البولية الرئيسية مجموعة صغيرة من معلومات التعريف حول الحيوان المانح، وذلك عند الكشف عن العضو الميكعي الأنفي للحيوان المتلقي. تنتمي البروتينات البولية الرئيسية إلى عائلة كبيرة من البروتينات المعروفة باسم الليبوكالينات. يُرمز إلى البروتينات البولية الرئيسية بواسطة مجموعة من الجينات، والتي تقع مجاورة لبعضها البعض في شريط واحد من الحمض النووي، والتي تختلف اختلافاً كبيرًا في عددها بين الأنواع: من 21 جين وظيفي على الأقل في الفئران إلى لا شيء في البشر. تشكل البروتينات البولية الرئيسية شكل قفاز مميز، تشمل بعض الروابط والمواد الكيميائية العضوية الصغيرة.
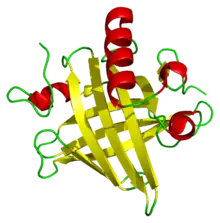
تم الإبلاغ لأول مرة عن البروتينات البولية في القوارض في عام 1932، خلال دراسات أجراها توماس أديس في سبب البيلة البروتينية. تعتبر البروتينات البولية الرئيسية من المواد المسببة للحساسية بقوة لدى البشر، كما أنها مسؤولة إلى حد كبير عن عدد من حالات الحساسية لدى الحيوانات، بما في ذلك القطط والخيول والقوارض. تظل وظيفة هذه البروتينات في الحيوانات غير معروفة، لكن ربما تشمل تنظيم استهلاك الطاقة. ومع ذلك، فبكونها بروتينات تفرزها الحيوانات قد تلعب أداء أدوار متعددة في الاتصال الكيميائي بين الحيوانات، كفيرومونات النقل والتثبيت في القوارض والخنازير. كما يمكن أن تلعب البروتينات البولية الرئيسية دور الفيرومونات مستقلةً بذاتها. أُثبت أن هذه البروتينات قد تحفز من النشاط العدواني لذكور الفئران، كام وُجد أن أحد هذه البروتينات الموجودة في بول ذكور الفئران تعمل كعامل جذب جنسي لإناثها. يمكن أن تلعب البروتينات البولية الرئيسية أيضا دورًا كإشارات بين الأنواع المختلفة: إذ تُظهر الفئران استجابة الخوف كعرض استجابي غريزي عند اكتشاف البروتينات البولية الرئيسية الخاصة بالحيوانات المفترسة مثل القطط والفئران.
الاكتشاف
يُخرج البشر الأصحاء بول خالٍ إلى حدٍ كبير من البروتينات. لذلك اهتم العلماء والأطباء بدراسة البيلة البروتينية منذ عام 1827، والتي تُعرّف بإفراز كميات زائدة من البروتينات في البول، وبالتالي تُعتبر بمثابة مؤشر لاعتلال الكلية .[1] ولفهمٍ أفضل لهذه الظاهرة، حاول العلماء دراسة هذه الظاهرة على حيوانات التجارب.[2] أشار العديد من العلماء في الفترة ما بين 1932 و 1933، بما في ذلك توماس أديس ، إلى النتائج المبهرة الخاصة بوجود البروتينات في بول بعض القوارض.[3][4][5] ومع ذلك، لم نحصل على أول وصف دقيق لبروتينات البول الرئيسية حتى ستينات القرن الماضي. وقد وجد أن البروتينات تصنع في المقام الأول عن طريق كبد الذكور وتُفرز عن طريق الكلى في البول بكميات كبيرة (بضعة ملليغرامات في اليوم الواحد).[6][7][8]
وحيث أن تلك البروتينات كانت قد سُميت بالفعل، ، تم العثور على إفراز هذه البروتينات من قبل الغدد المختلفة التي تخرج إفرازاتها مباشرةً إلى البيئة الخارجية. وتشمل هذه الغدد كل من الغدد الدمعية و النكافية و غدة تحت الفك السفلي و غدة تحت اللسان و القلفة و الغدد الثديية.[8][9][10] لا تُفرز بروتينات البول الرئيسية في بول بعض الأنواع مثل القطط و الخنازير ، بينما تُفرز بصورة أساسية في اللعاب.[11][12] يُستخدم مصطلح بروتينات البول الرئيسية البولية uMups في بعض الأحيان للتمييز بين البروتينات المُفرزة في البول وتلك الأخرى المُفرزة من الغدد الأخرى.[13]
جينات البروتينات البولية الرئيسية
اكتشف بين عامي 1979 و 1981 أن بروتينات البول الرئيسية تُخلّق بواسطة عائلة جينية تتألف من 15 و 35 من الجينات و الجينات الكاذبة في الفئران ونحو 20 جينًا في الجرزان.[14][15][16] كما تم تحديد عدد أكثر دقة من الجينات في مجموعة من الأنواع في عام 2008 من خلال تحليل تسلسل الحمض النووي للجينوم بأكمله.[17]
القوارض
يحتوي بول الجرزان على بروتينات بولية متماثلة؛ وعلى الرغم من أنها سُميت باسم مختلف، جلوبيولين- α2u إلا أنها عُرفت منذ ذلك الحين باسم بروتينات البول الرئيسية للجرزان.[19][20] تمتلك الجرزان 9 جينات و 13 جين كاذب لبروتينات البول الرئيسية تتجمع معًا عبر 1.1 قواعد عملاقة Megabases من الحمض النووي على كروموسوم 5. ومثلما حدث في الفئران، فقد تشكلت تلك الجينات بالإزدواج المتعدد. غير أن هذا حدث بشكل مستقل من الازدواجية في الفئران، وهذا يعني أن كلا من فصائل القوارض وسّعت عائلات جينات بروتينات البول الرئيسية خاصتها بشكل منفصل ، ولكن بالتوازي.[21]
غير القوارض
كان لدى الثدييات الأخرى التي تمت دراستها، بما في ذلك الخنازير والبقر والقطط والكلاب والجلاجو و قرد المكاك و الشمبانزي و إنسان الغاب جين واحد يرمز لبروتينات البول الرئيسية.إلا أن البعض كان لديهم أعداد أكبر: فالخيول كان لديها ثلاث جينات و الليمور الفأري الرمادي كان لديه على الأقل جينان. كما وُجد أن الحشرات و الأسماك و الزواحف والطيور والشقبانيات لديها تصاحب جيني مُعطل في مواضع كروموسومات جينات بروتينات البول الرئيسية، مما يرجح أن هذه العائلة من الجينات قد تكون خاصة بمشيمة الثدييات . لا يقتصر عدم امتلاك الإنسان على جينات بروتينات البول الرئيسية من بين الثدييات المشيمية؛ بل بالعكس فإنه يمتلك جين كاذب وحيد يحتوي على الطفرات التي تسببت في خلل في ربط الحمض النووي مما يجعل تلك البروتينات مختلة الوظيفة.
الوظيفة
بروتينات النقل
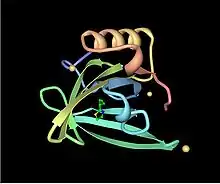
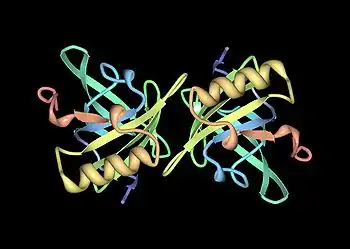
تُعتبر البروتينات البولية الرئيسية أحد أعضاء عائلة كبيرة من البروتينات منخفضة الوزن الجزيئي (~19 كيلو دالتون) تعرف باسم ا لليبوكالينات.[23] تمتلك هذه العائلة تركيب مميز من 8 أوراق بيتا مرتبة في طريقة عكس التوازي مع وجه واحد، مع لوالب ألفا عند كلا الطرفين. وبالتالي، فإنها تشكل شكل القفازات المميز، بما في ذلك جيب كوبي الشكل يربط بين الكيماويات العضوية الصغيرة بتقارب عالي.[24] يرتبط عدد من هذه الروابط بالبروتينات البولية الرئيسية الخاصة بالفئران، بما في ذلك 2-sec-butyl-4,5-dihydrothiazole (يختصر ك SBT or DHT), 6-hydroxy-6-methyl-3-heptanone (HMH) and 2,3 dihydro-exo-brevicomin (DHB).[25][25][26] أثبتت كل هذه الكيماويات المتواجدة بالبول أنها تلعب دور الفيرومونات— وهي إشارات جزيئية تفرز من قبل فرد واحد يؤدي إلى استجابة فطرية سلوكية في عضو آخر من نفس النوع.[27] أظهرت البروتينات البولية الرئيسية للفئران أنها تلعب أيضًا دور وظيفة مثبت الفيرومون، والتي توفر آلية بطء الانتشار مما يُقوي من فاعلية الفيرومونات الموجودة في رائحة بول الذكور.[28] ونظرًا لتنوع البروتينات البولية الرئيسية في القوارض ، كان يعتقد أصلاً أن مختلف البروتينات البولية الرئيسية تحتوي على جيوب ارتباط مختلفة التركيب وبالتالي ترتبط بمختلف الفيرومونات. بيد أن الدراسات التفصيلية وجدت أن معظم المواقع المتغيرة الموجودة على سطح البروتينات كان لها تأثير لا ُيذكر الارتباط بالروابط.[29]
ترتبط البروتينات البولية الرئيسية للفئران بروابط كيميائية صغيرة. يعتبر كلورو ديكان-1 هو الرابط الأكثر شيوعًا، بينما كان 2-ميثيل-N-فينيل-2-أكريلاميد وهكسا ديكان و2،6,11- ترايمثيل ديكان أقل شيوعًا .[30] ترتبط بروتينات الفئران الرئيسية أيضًا ب الليمونين-1 و 2-إيبوكسيد مما يؤدي إلى مرض الكلية فيما يسم باعتلال الكلية الزجاجي والذي سرعان ما يتطور إلى السرطان. ولاتُصاب الأنواع الأخرى بهذا المرض لعدم احتواء بولها على هذا النوع من الروابط الكيميائية بالتحديد.[31] وتبعًا لذلك، أصيبت الفئران المحورة جينيا بذلك المرض عند تعديل جيناتها للتعبير عن هذا النوع من البروتينات.[32] توجد البروتينات البولية الرئيسية ف الخنازير، والتي تسمى الليبوكالينات اللعابية لإفرازها من الغدد اللعابية، وترتبط بالأندروستينون و الأندروستينول والذي يلعب كلًا منهما دور الفيرومونات لاجتذاب اللإناث للتزاوج.
كشفت دراسات المعايرة الحرارية للسعرات أن البروتينات البولية الرئيسية والروابط المرتبطة بها (البايرازين[33][34][35][34] و الكحول [35][36] وا لثايازولين[37][38][39][38] و6-هيدروكسي-6-ميثيل-3-هيبتانون[39] وN-فينيلنفثيل أمين [40][41]) ظاهرة ارتباط غريبة. كان الموقع النشط لتلك البروتينات نادرًا ما يحتوي على الماء، مما يؤدي إلى ارتباط الروابط بواسطة قوى التشتت الحراري. وهذا يتعارض مع معظم البروتينات الأخرى، والتي عادةً ما تستهلك القوى الحرارية الناتجة عن تحريك جزيئات الماء. سُميت تلك الظاهرة الغريبة ب "التأثير الكاره للماء الغير اعتيادي."[41]
الفيرومونات
سعت الدراسات للكشف عن الوظيفة الدقيقة لبروتينات البول الرئيسية في الاتصال الفيروموني. وُجد أن البروتينات البولية الرئيسية تُشجع البلوغ وتسرع من دورة الشبق عند إناث الفئران، بما في ذلك تأثير فاندينبيرغ و يتون.[35][42] بيد أنه وُجد أن البروتينات البولية الرئيسية التي تجذب الإناث تحتاج إلى الذوبان في بول الذكور، مما يشير إلى أن تلك البروتينات تحتاج إلى البول لكي تؤدي وظيفتها . وُجد في عام 2007 أن البروتينات البولية الرئيسية الموجودة عادةً في بول ذكور الفئران تُخلق في البكتيريا المحورة وراثيا، ، وبالتالي فإنها تخلو عادةً من المواد الكيميائية التي ترتبط بها عادةً. كما وُجد أن هذه البروتينات كانت كافية لتعزيز سلوك العدوانية في الذكور، حتى في غياب البول. بالإضافة إلى ذلك، تم العثور على أن البروتينات البولية المُخلّقة في البكتيريا يمكن أن تُنشط الخلايا العصبية الحسية الشمية في الجهاز الميكعي الأنفي VNO، وهو نظام فرعي من الأنف لديه قدرة الكشف عن الفيرومونات عبر مستقبلات حسية محددة، لدى الفئران والجرذان.[17][43] ويدلل هذا على أن البروتينات البولية الرئيسية يمكن أن تلعب دور الفيرومونات بنفسها مستقلة عن الروابط الخاصة بها.[44]
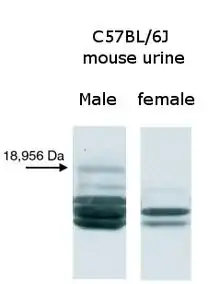
اتساقًا مع دور تلك البروتينات في السلوك العدواني بين ذكور الفئران، تفرز ذكور الفئران البالغين بروتينات بولية رئيسية أكثر بشكل ملحوظ في بولهم عن الإناث أو الأحداث أو ذكور الفئران المخصيين . الآلية الدقيقة التي تقود لهذ الفرق بين الجنسن أمر معقد، لكن على الأقل توجد ثلاثة هرمونات – التستوستيرون و هرمون النمو و هرمون الغدة الدرقية – معروفة بتأثيرها إيجابيًا على إنتاج البروتينات البولية الرئيسية في الفئران.[44][45] يحتوي بول فئران المنازل على تركيبات متغيرة من أربع إلى سبع بروتينات رئيسية مُميزة للفأر الواحد. وُجد أن سلالات الفئران المخبرية الفطرية مثل BALB/c و C57BL/6 تُفرز بروتينات مختلفة في بولها . ومع ذلك، على عكس فئران المنازل، يفرز الأفراد المختلفين من نفس السلالة نفس النمط البروتيني، والذي يُعد قطعة أثرية من أجيال عديدة من التوالد الداخلي.[46][47] يختلف هذا النوع من البروتينات البولية الرئيسية عن الأنواع الأخرى: فمن الممكن أن تُنتجه نسبة كبيرة من ذكور الفئران ولا تنتجه إناثها بأي حال من الأحوال. عند تخليق هذا النوع من البروتين في البكثيريا واستخدامه في اختبارات السلوك، وُجد أنه يجذب الإناث.تم اختبار الأنواع الأخرى من تلك البروتينات ولم تتحلى بنفس مواصفات الجذب مما يُرجح من تفرد البروتينات البولية الرئيسية للذكور في كونها فيرومونات جنسية.سمّى العلماء هذا النوع من الفيرومونات دارسين Darcin في دلالة مرحة إلى فيتزويليام دارسي البطل الرومانسي لرواية كبرياء وتحامل.[48][49] بأخذ كل هذا في الاعتبار، توفر تلك الأنماط المعقدة من البروتينات البولية الريسية طائفة من المعلومات خول الحيوانات المانحة، مثل الجنس والخصوبة والهيمنة الاجتماعية و العمر و التنوع الوراثي والقرابة.[50][51] تمتلك الفئران البرية (على عكس الفئران المختبرية التي تتطابق وراثيًا ولذلك توفر أنماط مماثلة من البروتينات البولية الرئيسية) أنماط فريدة من البروتينات البولية الرئيسية التي تعمل بمثابة "رمز شريطي" يحدد بدقة صاحب تلك الرائحة.
توفر مجموعة جينات البروتينات البولية الرئيسية في فئران المنازل إشارة رائحة متعددة الأشكال للغاية ذات بصمة وراثية واضحة. أظهرت الفئران البرية المُرباة في مرفقات شبه طبيعبة تجنب التوالد الداخلي. أسفر هذا التجنب عن نقص كبير في التزاوج الناجح بين الفئران التي تتشارك نسخًا متنوعة من البروتينات البولية الرئيسية( مما أنتج تطابق تام).[52]
الكيرومونات
بالإضافة إلى لعب البروتينات البولية الرئيسية دور الإشارات الاجتماعية بين أفراد نفس النوع، تستطيع أيضًا أن تلعب دور الكيرومونات- وهي إشارات كيميائية تنقل المعلومات بين الأنواع.[53][54][55] تخاف الفئران بشكل غريزي من رائحة الحيوانات التي تفترسها، بما في ذلك القطط والجرزان. يحدث هذا أيضًا في الفئران المخبرية التي تم عزلها عن الحيوانات المفترسة لمئات الأجيال.[56] تم التعرف على إشارتين للبروتينات المتماثلة عند فصل وتنقية الإشارات الكيميائية المسؤلة عن استجابة الخوف من بول الجرزان ولعاب القطط : Fel d 4 (Felis domesticus allergen 4) المنتج من جينات البروتينات البولية الرئيسية الخاصة بالقطط، Rat n 1 (Rattus norvegicus allergen 1) المنتج من جينات البروتينات البولية الرئيسية الخاصة بالفئران تخاف الفئران أيضًا من هذه البروتينات حتى عندما تكون مصنوعة في البكتيريا، بينما لم تظهر الحيوانات المحورة جينيًا معدومة القدرة لاكتشاف تلك البروتينات أي خوفٍ من الفئران، مما يدل على أهميتها في ابتداء سلوك الخوف.[53][57] تظل كيفية ابداء السلوكات المتباينة نتيجة التعرف على هذه البروتينات غير واضحة، لكن أظهرت البروتينات البولية الرئيسية للفئران والحيوانات المفترسة قدرتها على تنشيط أنماط مختلفة من الأعصاب الحسية للفئران المستقبلة، من خلال دوائر عصبية مختلفة.[53][54] وتظل مستقبلات الفيرومونات المسؤولة عن اكتشاف البروتينات البولية الرئيسية غير معروفة، لذا يُظن أن تكون تلك أحد عناصر عائلة V2R.[17][54]
المواد المسببة للحساسية
تُعتبر البروتينات البولية الرئيسية باعتبارها أحد عناصر عائلة الليبوكالينات من المواد المسببة للحساسية بقوة لدى البشر.[59] يظل السبب لهذا غير معروف، إلا أنه يمكن أخذ التابه الجزيئي بين هذه البروتينات والليبوكالينات في الاعتبار.[60] تعد البروتينات الناتجة عن جينات Mup17 والمعروفة ك Mus m 1, Ag1 or MA1 مسؤولة بشكل كبير عن التسبب في حساسية البشر لبول الفئران .[61][62] تظل تلك البروتينات في حالة مستقرة في البيئة الخارجية، فقد وجدت إحدى الدراسات أن 95% من المنازل الداخية و 82% من جميع المنازل بالولايات المتحدة تحتوي على مستويات قابلة للكشف عنها في كل حجرة على الأقل..[63][64] كما وجدت إحدى الدراسات بالولايات المتحدة تلك الحساسية في 33% من المنازل الداخلية و21% من جميع المنازل.[65] يعتبر التعرض لبروتينات البول الريسية للفئران أحد عوامل الخطر للإصابة بالربو في الصغر وأحد العوامل الرئيسية للإصابة باحساسية لحيوانات المختبر، وهو مرض مهني يُصاب به التقنيين والباحثين في مختبرات الحيوان.[66][67][68][69] One study found that two-thirds of laboratory workers who had developed asthmatic reactions to animals had جسم مضاد to Rat n 1.[70]
الأيض
يظل الدور الوظيفي للبروتينات البولية الرئيسية أقل وضوحًا في بعض الحيوانات المُنتجة له، في حين تم دراسة هذا الدور جيدًا في الحيوانات التي تقو بإخراجه. ومع ذلك، تم الافتراض في عام 2009 أن البروتينات البولية الرئيسية Mups قد تكون مرتبطة مع تنظيم استهلاك الطاقة في الفئران. وجد العلماء أن الفئران المعدلة وراثيًا السمينة والمصابة بداء السكري تنتج الحمض النووي الريبوزي الخاص ببروتينات البول الرئيسية Mup RNA ثلاثين مرة أقل من أشقائها الهزلاء.[71] عندما حُقنت تلك البروتينات مباشرةً في مجرى دم هذه الفئران، لوحظت زيادة في استهلاك الطاقة والنشاط البدني و درجة حرارة الجسم وانخفاض مماثل في اختلال تحمل الجلوكوز و مقاومة الأنسولين. ويُقترح أن تحدث تلك الآثار المفيدة في أيض الطاقة من خلال تعزيز وظيفة الميتوكوندريا في العضلات الهيكلية. وجدت دراسة أخرى انخفاض نسبة البروتينات البولية الرئيسية الفئران السمينة بسبب النظام الغذائي. يُعتبر وجود هذه البروتينات في مجرى دم هذه الفئران في هذه الحالة عاملًا مثبطًا لإنتاج الجلوكوز من خلال تثبيط التعبير الجيني في الكبد.[72]
مراجع
- Comper WD, Hilliard LM, Nikolic-Paterson DJ, Russo LM (2008). "Disease-dependent mechanisms of albuminuria". Am. J. Physiol. Renal Physiol. 295 (6): F1589–600. doi:10.1152/ajprenal.00142.2008. PMID 18579704. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lemley KV, Pauling L (1994). "Thomas Addis: 1881–1949". Biographical Memoirs, National Academy of Sciences. 63: 1–46. مؤرشف من الأصل في 8 مايو 2014. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Addis T. (1932). "Proteinuria and cylinduria". Proceedings of the California Academy of Sciences. 2: 38–52. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bell ME. (1933). "Albuminuria in the normal male rat". Journal of Physiology. 79 (2): 191–93. PMC 1394952. PMID 16994453. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Parfentjev IA, Perlzweig WA (1933). "The Composition of the Urine of White Mice". The Journal of Biological Chemistry. 100 (2): 551–55. مؤرشف من الأصل في 13 فبراير 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Finlayson JS, Asofsky R, Potter M, Runner CC (1965). "Major urinary protein complex of normal mice: origin". Science. 149 (3687): 981–82. doi:10.1126/science.149.3687.981. PMID 5827345. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Roy AK, Neuhaus OW (1966). "Identification of rat urinary proteins by zone and immunoelectrophoresis". Proceedings of the Society for Experimental Biology and Medicine. 121 (3): 894–99. doi:10.3181/00379727-121-30917. PMID 4160706. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Roy AK, Neuhaus OW (1966). "Proof of the hepatic synthesis of a sex-dependent protein in the rat". Biochimica et Biophysica Acta. 127 (1): 82–87. doi:10.1016/0304-4165(66)90478-8. PMID 4165835. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Gubits RM, Lynch KR, Kulkarni AB, et al. (1984). "Differential regulation of alpha 2u globulin gene expression in liver, lachrymal gland, and salivary gland". The Journal of Biological Chemistry. 259 (20): 12803–809. PMID 6208189. مؤرشف من الأصل في 11 مايو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Shahan K, Denaro M, Gilmartin M, Shi Y, Derman E (1987). "Expression of six mouse major urinary protein genes in the mammary, parotid, sublingual, submaxillary, and lachrymal glands and in the liver". Molecular and Cellular Biology. 7 (5): 1947–54. PMC 365300. PMID 3600653. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Smith W, Butler AJ, Hazell LA, et al. (2004). "Fel d 4, a cat lipocalin allergen". Clinical and Experimental Allergy. 34 (11): 1732–38. doi:10.1111/j.1365-2222.2004.02090.x. PMID 15544598. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Loebel D, Scaloni A, Paolini S, et al. (2000). "Cloning, post-translational modifications, heterologous expression and ligand-binding of boar salivary lipocalin". The Biochemical Journal. 350 (Pt 2): 369–79. doi:10.1042/0264-6021:3500369. PMC 1221263. PMID 10947950. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Beynon RJ, Hurst JL (2003). "Multiple roles of major urinary proteins in the house mouse, Mus domesticus". Biochemical Society Transactions. 31 (Pt 1): 142–46. doi:10.1042/BST0310142. PMID 12546672. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Kurtz DT (1981). "Rat alpha 2u globulin is encoded by a multigene family". Journal of Molecular and Applied Genetics. 1 (1): 29–38. PMID 6180115. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Hastie ND, Held WA, Toole JJ (1979). "Multiple genes coding for the androgen-regulated major urinary proteins of the mouse". Cell. 17 (2): 449–57. doi:10.1016/0092-8674(79)90171-5. PMID 88267. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bishop JO, Clark AJ, Clissold PM, Hainey S, Francke U (1982). "Two main groups of mouse major urinary protein genes, both largely located on chromosome 4". The EMBO Journal. 1 (5): 615–20. PMC 553096. PMID 6329695. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Chamero P, Marton TF, Logan DW, et al. (2007). "Identification of protein pheromones that promote aggressive behaviour". Nature. 450 (7171): 899–902. doi:10.1038/nature05997. PMID 18064011. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Mudge JM, Armstrong SD, McLaren K, et al. (2008). "Dynamic instability of the major urinary protein gene family revealed by genomic and phenotypic comparisons between C57 and 129 strain mice". Genome Biology. 9 (5): R91. doi:10.1186/gb-2008-9-5-r91. PMC 2441477. PMID 18507838. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Hurst J, Beynon RJ, Roberts SC, Wyatt TD (2007). Urinary Lipocalins in Rodenta:is there a Generic Model?. Springer New York. ISBN 978-0-387-73944-1. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cavaggioni A, Mucignat-Caretta C (2000). "Major urinary proteins, alpha(2U)-globulins and aphrodisin". Biochimica et Biophysica Acta. 1482 (1–2): 218–28. doi:10.1016/S0167-4838(00)00149-7. PMID 11058763. الوسيط
|CitationClass=تم تجاهله (مساعدة) - McFadyen DA, Addison W, Locke J (1999). "Genomic organization of the rat alpha 2u-globulin gene cluster". Mammalian Genome. 10 (5): 463–70. doi:10.1007/s003359901024. PMID 10337619. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Böcskei Z, Groom CR, Flower DR, et al. (1992). "Pheromone binding to two rodent urinary proteins revealed by X-ray crystallography". Nature. 360 (6400): 186–88. doi:10.1038/360186a0. PMID 1279439. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Flower DR (1996). "The lipocalin protein family: structure and function". The Biochemical Journal. 318 (1): 1–14. PMC 1217580. PMID 8761444. مؤرشف من الأصل في 22 أكتوبر 2002. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ganfornina MD, Gutiérrez G, Bastiani M, Sánchez D (2000). "A phylogenetic analysis of the lipocalin protein family". Molecular Biology and Evolution. 17 (1): 114–26. doi:10.1093/oxfordjournals.molbev.a026224. PMID 10666711. مؤرشف من الأصل في 7 يوليو 2009. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Halpern M, Martínez-Marcos A (2003). "Structure and function of the vomeronasal system: an update". Progress in Neurobiology. 70 (3): 245–318. doi:10.1016/S0301-0082(03)00103-5. PMID 12951145. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Armstrong SD, Robertson DH, Cheetham SA, Hurst JL, Beynon RJ (2005). "Structural and functional differences in isoforms of mouse major urinary proteins: a male-specific protein that preferentially binds a male pheromone". The Biochemical Journal. 391 (Pt 2): 343–50. doi:10.1042/BJ20050404. PMC 1276933. PMID 15934926. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Stowers L, Marton TF (2005). "What is a pheromone? Mammalian pheromones reconsidered". Neuron. 46 (5): 699–702. doi:10.1016/j.neuron.2005.04.032. PMID 15924856. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Hurst JL, Robertson DH, Tolladay U, Beynon RJ (1998). "Proteins in urine scent marks of male house mice extend the longevity of olfactory signals". Animal Behaviour. 55 (5): 1289–97. doi:10.1006/anbe.1997.0650. PMID 9632512. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Darwish Marie A, Veggerby C, Robertson DH, et al. (2001). "Effect of polymorphisms on ligand binding by mouse major urinary proteins". Protein Science. 10 (2): 411–17. doi:10.1110/ps.31701. PMC 2373947. PMID 11266626. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Rajkumar R, Ilayaraja R, Mucignat C, Cavaggioni A, Archunan G (2009). "Identification of alpha2u-globulin and bound volatiles in the Indian common house rat (Rattus rattus)". Indian Journal of Biochemistry & Biophysics. 46 (4): 319–24. PMID 19788064. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lehman-McKeeman LD, Caudill D (1992). "Biochemical basis for mouse resistance to hyaline droplet nephropathy: lack of relevance of the alpha 2u-globulin protein superfamily in this male rat-specific syndrome". Toxicology and Applied Pharmacology. 112 (2): 214–21. doi:10.1016/0041-008X(92)90190-4. PMID 1371614. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lehman-McKeeman LD, Caudill D (1994). "d-Limonene induced hyaline droplet nephropathy in alpha 2u-globulin transgenic mice". Fundamental and Applied Toxicology. 23 (4): 562–68. doi:10.1006/faat.1994.1141. PMID 7532604. الوسيط
|CitationClass=تم تجاهله (مساعدة) - [[بايرازين|Bingham, R.; et al. (2004). "Thermodynamics of binding of 2-methoxy-3-isopropylpyrazine and 2-methoxy-3-isobutylpyrazine to the major urinary protein". J. Am. Chem. Soc. 126 (6): 1675–1681. doi:10.1021/ja038461i. PMID 14871097. الوسيط
|CitationClass=تم تجاهله (مساعدة)]] - [[بايرازين|Barratt, E.; et al. (2005). "Van der Waals interactions dominate ligand-protein association in a protein binding site occluded from solvent water". J. Am. Chem. Soc. 127 (33): 11827–11834. doi:10.1021/ja0527525. PMID 16104761. الوسيط
|CitationClass=تم تجاهله (مساعدة)]] - Mucignat-Caretta C, Caretta A, Cavaggioni A (1995). "Acceleration of puberty onset in female mice by male urinary proteins". J. Physiol. (Lond.). (Pt 2): 517–22. PMC 1156539. PMID 7473215. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Malham, R.; et al. (2005). "Strong solute-solute dispersive interactions in a protein-ligand complex". J. Am. Chem. Soc. 127 (48): 17061–17067. doi:10.1021/ja055454g. PMID 16316253. الوسيط
|CitationClass=تم تجاهله (مساعدة) - [[ثيازوليدينديون|Sharrow, S.D.; Novotny, M.V.; Stone, M. J. (2003). "Thermodynamic analysis of binding between mouse major urinary protein-I and the pheromone 2-sec- butyl-4,5-dihydrothiazole". Biochemistry. 42 (20): 6302–6309. doi:10.1021/bi026423q. PMID 12755635. الوسيط
|CitationClass=تم تجاهله (مساعدة)]] - [[ثيازوليدينديون|Timm, D.E.; Baker, L.J.; Mueller, H.; Zidek, L.; Novotny, M.V. (2001). "Structural basis of pheromone binding to mouse major urinary protein (MUP-I)". Protein Sci. 10: 997–1004. doi:10.1110/ps.52201. PMC 2374202. PMID 11316880. الوسيط
|CitationClass=تم تجاهله (مساعدة)]] - Sharrow, S. D.; et al. (2005). "Thermodynamic consequences of disrupting a water-mediated hydrogen bond network in a protein: pheromone complex". Protein Sci. 14 (1): 249–256. doi:10.1110/ps.04912605. PMC 2253314. PMID 15608125. الوسيط
|CitationClass=تم تجاهله (مساعدة) - T. A; et al. (2009). "The binding cavity of mouse major urinary protein is optimised for a variety of ligand binding modes". Biochem. Bioph. Res. Co. 390 (4): 1266–1271. doi:10.1016/j.bbrc.2009.10.133. PMID 19878650. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Homans, S. W. (2007). "Water, water everywhere — except where it matters?". Drug Discovery Today. 12 (13–14): 534–539. doi:10.1016/j.drudis.2007.05.004. PMID 17631247. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Marchlewska-koj A, Caretta A, Mucignat-Caretta C, Olejniczak, P (2000). "Stimulation of estrus in female mice by male urinary proteins". Journal of Chemical Ecology. 26 (10): 2355–65. doi:10.1023/A:1005578911652. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Krieger J, Schmitt A, Löbel D, et al. (1999). "Selective activation of G protein subtypes in the vomeronasal organ upon stimulation with urine-derived compounds". The Journal of Biological Chemistry. 274 (8): 4655–62. doi:10.1074/jbc.274.8.4655. PMID 9988702. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Aggression protein found in mice". بي بي سي نيوز. 5 December 2007. مؤرشف من الأصل في 1 مارس 2019. اطلع عليه بتاريخ 26 سبتمبر 2009. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Robertson DH, Hurst JL, Bolgar MS, Gaskell SJ, Beynon RJ (1997). "Molecular heterogeneity of urinary proteins in wild house mouse populations". Rapid Communications in Mass Spectrometry. 11 (7): 786–90. doi:10.1002/(SICI)1097-0231(19970422)11:7<786::AID-RCM876>3.0.CO;2-8. PMID 9161047. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Robertson DH, Cox KA, Gaskell SJ, Evershed RP, Beynon RJ (1996). "Molecular heterogeneity in the Major Urinary Proteins of the house mouse Mus musculus". The Biochemical Journal. 316 (Pt 1): 265–72. PMC 1217333. PMID 8645216. مؤرشف من الأصل في 6 مايو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cheetham SA, Smith AL, Armstrong SD, Beynon RJ, Hurst JL (2009). "Limited variation in the major urinary proteins of laboratory mice". Physiology & Behavior. 96 (2): 253–61. doi:10.1016/j.physbeh.2008.10.005. PMID 18973768. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Roberts SA, Simpson DM, Armstrong SD, et al. (2010). "Darcin: a male pheromone that stimulates female memory and sexual attraction to an individual male's odour". BMC Biol. 8 (1): 75. doi:10.1186/1741-7007-8-75. PMC 2890510. PMID 20525243. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Moskowitz, C (3 June 2010). "Biologists Learn Why Mice Go Gaga for Urine". FoxNews.com. FOX News Network. مؤرشف من الأصل في 24 سبتمبر 2015. اطلع عليه بتاريخ 09 يونيو 2010. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Hurst JL, Payne CE, Nevison CM, et al. (2001). "Individual recognition in mice mediated by major urinary proteins". Nature. 414 (6864): 631–34. doi:10.1038/414631a. PMID 11740558. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Thom MD, Stockley P, Jury F, Ollier WE, Beynon RJ, Hurst JL (2008). "The direct assessment of genetic heterozygosity through scent in the mouse". Current Biology. 18 (8): 619–23. doi:10.1016/j.cub.2008.03.056. PMID 18424142. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Sherborne AL, Thom MD, Paterson S, Jury F, Ollier WE, Stockley P, Beynon RJ, Hurst JL (2007). "The genetic basis of inbreeding avoidance in house mice". Curr Biol. 17 (23): 2061–2066. doi:10.1016/j.cub.2007.10.041. PMC 2148465. PMID 17997307. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Papes F, Logan DW, Stowers L (2010). "The Vomeronasal Organ Mediates Interspecies Defensive Behaviors through Detection of Protein Pheromone Homologs". Cell. 141 (4): 692–703. doi:10.1016/j.cell.2010.03.037. PMC 2873972. PMID 20478258. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Rodriguez I (2010). "The chemical MUPpeteer". Cell. 141 (4): 568–70. doi:10.1016/j.cell.2010.04.032. PMID 20478249. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Why mice fear the smell of cats". بي بي سي نيوز. 17 May 2010. مؤرشف من الأصل في 11 مايو 2020. اطلع عليه بتاريخ 18 مايو 2010. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ehrenberg, Rachel (5 June 2010). "Fight or flee, it's in the pee". Science News. مؤرشف من الأصل في 12 أكتوبر 2012. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bhanoo, Sindya (17 May 2010). "When a Mouse Smells a Rat". The New York Times. مؤرشف من الأصل في 10 فبراير 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lascombe MB, Grégoire C, Poncet P, et al. (2000). "Crystal structure of the allergen Equ c 1. A dimeric lipocalin with restricted IgE-reactive epitopes". The Journal of Biological Chemistry. 275 (28): 21572–77. doi:10.1074/jbc.M002854200. PMID 10787420. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lockey R, Ledford DK (2008). "Mammalian Allergens". Allergens and Allergen Immunotherapy. Informa Health Care. صفحات 201–218. ISBN 978-1-4200-6197-0. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Virtanen T, Zeiler T, Mäntyjärvi R (1999). "Important animal allergens are lipocalin proteins: why are they allergenic?". Int. Arch. Allergy Immunol. 120 (4): 247–58. doi:10.1159/000024277. PMID 10640908. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Logan DW, Marton TF, Stowers L (2008). "Species specificity in major urinary proteins by parallel evolution". PLoS ONE. 3 (9): e3280. doi:10.1371/journal.pone.0003280. PMC 2533699. PMID 18815613. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lorusso JR, Moffat S, Ohman JL (1986). "Immunologic and biochemical properties of the major mouse urinary allergen (Mus m 1)". The Journal of Allergy and Clinical Immunology. 78 (5 Pt 1): 928–37. doi:10.1016/0091-6749(86)90242-3. PMID 3097107. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cohn RD, Arbes SJ, Yin M, Jaramillo R, Zeldin DC (2004). "National prevalence and exposure risk for mouse allergen in US households". The Journal of Allergy and Clinical Immunology. 113 (6): 1167–71. doi:10.1016/j.jaci.2003.12.592. PMID 15208600. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Phipatanakul W, Eggleston PA, Wright EC, Wood RA (2000). "Mouse allergen. I. The prevalence of mouse allergen in inner-city homes. The National Cooperative Inner-City Asthma Study". The Journal of Allergy and Clinical Immunology. 106 (6): 1070–74. doi:10.1067/mai.2000.110796. PMID 11112888. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Perry T, Matsui E, Merriman B, Duong T, Eggleston P (2003). "The prevalence of rat allergen in inner-city homes and its relationship to sensitization and asthma morbidity". The Journal of Allergy and Clinical Immunology. 112 (2): 346–52. doi:10.1067/mai.2003.1640. PMID 12897741. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wood RA (2001). "Laboratory animal allergens". ILAR Journal. 42 (1): 12–16. doi:10.1093/ilar.42.1.12. PMID 11123185. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Gaffin JM, Phipatanakul W (2009). "The role of indoor allergens in the development of asthma". Current Opinion in Allergy and Clinical Immunology. 9 (2): 128–35. doi:10.1097/ACI.0b013e32832678b0. PMC 2674017. PMID 19326507. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Pongracic JA, Visness CM, Gruchalla RS, Evans R, Mitchell HE (2008). "Effect of mouse allergen and rodent environmental intervention on asthma in inner-city children". Annals of Allergy, Asthma & Immunology. 101 (1): 35–41. doi:10.1016/S1081-1206(10)60832-0. PMID 18681082. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Gordon S, Preece R (2003). "Prevention of laboratory animal allergy". Occupational Medicine. 53 (6): 371–77. doi:10.1093/occmed/kqg117. PMID 14514903. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Platts-Mills TA, Longbottom J, Edwards J, Cockroft A, Wilkins S (1987). "Occupational asthma and rhinitis related to laboratory rats: serum IgG and IgE antibodies to the rat urinary allergen". The Journal of Allergy and Clinical Immunology. 79 (3): 505–15. doi:10.1016/0091-6749(87)90369-1. PMID 3819230. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Hui X, Zhu W, Wang Y, et al. (2009). "Major urinary protein-1 increases energy expenditure and improves glucose intolerance through enhancing mitochondrial function in skeletal muscle of diabetic mice". The Journal of Biological Chemistry. 284 (21): 14050–57. doi:10.1074/jbc.M109.001107. PMC 2682853. PMID 19336396. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Zhou Y, Jiang L, Rui L (2009). "Identification of MUP1 as a regulator for glucose and lipid metabolism in mice". The Journal of Biological Chemistry. 284 (17): 11152–59. doi:10.1074/jbc.M900754200. PMC 2670120. PMID 19258313. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة علم الأحياء
بوابة علم الأحياء بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية
