عين (تشريح)
العين (بالإنجليزية: Eye) هي أعضاء في الجهاز البصري . إنها تزود الحيوانات بالرؤية والقدرة على تلقي التفاصيل المرئية ومعالجتها، فضلاً عن تمكين العديد من وظائف الاستجابة للصور المستقلة عن الرؤية. تكتشف العيون الضوء وتحوله إلى نبضات كهربائية كيميائية في الخلايا العصبية . في الكائنات الحية العليا، تعتبر العين نظامًا بصريًا معقدًا يجمع الضوء من البيئة المحيطة، وينظم شدته من خلال غشاء، ويركزها من خلال تجميع قابل للتعديل من العدسات لتكوين صورة، ويحول هذه الصورة إلى مجموعة من الإشارات الكهربائية، وينقل هذه الإشارات إلى الدماغ من خلال مسارات عصبية معقدة تربط العين عبر العصب البصري بالقشرة البصرية ومناطق أخرى من الدماغ. تأتي العيون ذات القدرة على التحليل في عشرة أشكال مختلفة اختلافًا جوهريًا، و 96٪ من أنواع الحيوانات تمتلك نظامًا بصريًا معقدًا.[1] عيون حل صورة موجودة في الرخويات، حبليات والمفصليات .[2]
| عين (تشريح) | |
|---|---|
 | |
| تفاصيل | |
| يتكون من | غرفة العين الخلفية |
| نوع من | عضو حيواني |
| ترمينولوجيا أناتوميكا | 15.2.00.001 و A01.1.00.007 |
| FMA | 54448 |
| UBERON ID | 0000970 |
| ن.ف.م.ط. | A01.456.505.420، وA09.371 |
| ن.ف.م.ط. | D005123 |


العين هي شبكة كروية وقطر عين الإنسان البالغ يصل إلى 2.5 سم تقع العين في مقدمة الجمجمة محمية داخل محجر العظمي، والعين قادرة على التحرك داخل المحجر بصورة حرة بمساعدة جهاز معقد من العضلات الطولية والعرضية المحيطة بالعين داخل المحجر.و في منطقة الحاجبين يبرز عظم الجمجمة لحماية العين من الاصابات الخارجية ومعروف ان للعين 6 عضلات خاصة تحركها.
العين هي عضو يلتقط الضوء الذي تعكسه الأشياء، وهي المسؤولة عن الإبصار.[3] وهي عبارة عن مستقبل حسي يتفاعل مع الضوء الصادر من الشيء المريء أو المنعكس منه ويقوم عضو(الشبكية) في العين بتحويل هدا الضوء لسيالة عصبية التي تنقل عبر الأعصاب ومن تم نحو الباحة البصرية الموجودة في الدماغ.
أبسط العيون، عيون الحفرة، هي بقع العين التي يمكن وضعها في حفرة لتقليل زوايا الضوء التي تدخل وتؤثر في بقعة العين، للسماح للكائن الحي باستنتاج زاوية الضوء الوارد.[1] من العيون الأكثر تعقيدًا، ترسل الخلايا العقدية الحساسة للضوء في شبكية العين إشارات على طول السبيل الشبكي الوهمي إلى النوى فوق التصالبية لإحداث تعديل الساعة البيولوجية وإلى المنطقة أمام المستقيم للتحكم في انعكاس الضوء الحدقي .
نظرة عامة

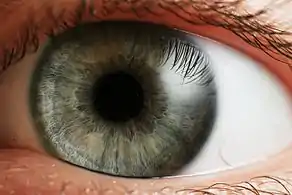
يمكن للعيون المعقدة أن تميز الأشكال والألوان . تتضمن المجالات المرئية للعديد من الكائنات الحية، وخاصة الحيوانات المفترسة، مساحات كبيرة من الرؤية ثنائية العين لتحسين إدراك العمق . في الكائنات الحية الأخرى، يتم وضع العيون من أجل زيادة مجال الرؤية، مثل الأرانب والخيول، التي لديها رؤية أحادية .
تطورت العيون البدائية الأولى بين الحيوانات 600 حول وقت الانفجار الكمبري .[4] آخر سلف مشترك من الحيوانات تمتلك مجموعة أدوات الكيميائية الحيوية اللازمة لرؤية، وعيون أكثر تقدما تطورت في 96٪ من الأنواع الحيوانية في ستة من ~ 35 [lower-alpha 1] الرئيسي من الكائنات الحية .[1] في معظم الفقاريات وبعض الرخويات، تعمل العين عن طريق السماح للضوء بالدخول والإسقاط على لوحة من الخلايا الحساسة للضوء، والمعروفة باسم شبكية العين، في مؤخرة العين. تقوم الخلايا المخروطية (للون) وخلايا القضيب (للتباين في الإضاءة المنخفضة) في الشبكية باكتشاف الضوء وتحويله إلى إشارات عصبية من أجل الرؤية. ثم يتم نقل الإشارات البصرية إلى الدماغ عبر العصب البصري . عادة ما تكون هذه العيون كروية تقريبًا، مليئة بمادة شفافة تشبه الهلام تسمى الخلط الزجاجي، مع عدسة تركيز وقزحية في كثير من الأحيان ؛ يؤدي ارتخاء أو شد العضلات حول القزحية إلى تغيير حجم بؤبؤ العين، وبالتالي تنظيم كمية الضوء التي تدخل العين، [5] وتقليل الانحرافات عند وجود ضوء كافٍ.[6] تتميز عيون معظم رأسيات الأرجل والأسماك والبرمائيات والثعابين بأشكال عدسات ثابتة، ويتم تحقيق التركيز البؤري عن طريق تصغير العدسة - على غرار طريقة تركيز الكاميرا .[7]
تم العثور على العيون المركبة بين المفصليات وتتكون من العديد من الجوانب البسيطة التي، اعتمادًا على تفاصيل التشريح، قد تعطي إما صورة واحدة أو صور متعددة، لكل عين. كل مستشعر له عدسته الخاصة وخلية (خلايا) حساسة للضوء. تحتوي بعض العيون على ما يصل إلى 28000 مستشعر من هذا القبيل، مرتبة بشكل سداسي، والتي يمكن أن توفر مجال رؤية كامل بمقدار 360 درجة. تركيب العين حساس للحركة. بعض المفصليات، بما في ذلك العديد من ملتويات الأجنحة، لها عيون مركبة من جوانب قليلة فقط، ولكل منها شبكية قادرة على تكوين صورة، وخلق الرؤية. مع كل عين ترى شيئًا مختلفًا، يتم إنتاج صورة مدمجة من جميع العيون في الدماغ، مما يوفر صورًا مختلفة جدًا وعالية الدقة.
يمتلك جمبري السرعوف رؤية ألوان فائقة الطيفية التفصيلية، وقد تم الإبلاغ عن أنه يمتلك أكثر أنظمة رؤية الألوان تعقيدًا في العالم.[8] ثلاثية الفصوص، التي انقرضت الآن، لها عيون مركبة فريدة من نوعها. استخدموا بلورات الكالسيت الصافية لتشكيل عدسات عيونهم. في هذا، تختلف عن معظم المفصليات الأخرى التي لها عيون ناعمة. اختلف عدد العدسات في مثل هذه العين ؛ ومع ذلك، كان لبعض ثلاثية الفصوص واحدة فقط، والبعض الآخر لديه آلاف العدسات في عين واحدة.
على عكس العيون المركبة، فإن العيون البسيطة هي تلك التي تحتوي على عدسة واحدة. على سبيل المثال، تحتوي العناكب القافزة على زوج كبير من العيون البسيطة ذات مجال رؤية ضيق، تدعمها مجموعة من العيون الأخرى الأصغر للرؤية المحيطية . تمتلك بعض يرقات الحشرات، مثل اليرقات، نوعًا مختلفًا من العين البسيطة ( الجذعية ) التي عادةً ما توفر صورة تقريبية فقط، ولكن (كما في يرقات المنشار) يمكن أن تمتلك قوى حل تبلغ 4 درجات من القوس، وتكون حساسة للاستقطاب وقادرة على الزيادة حساسيته المطلقة في الليل بعامل 1000 أو أكثر.[9] يمكن العثور على بعض أبسط العيون، التي تسمى عين بسيطة، في الحيوانات مثل بعض القواقع، والتي لا تستطيع في الواقع "الرؤية" بالمعنى الطبيعي. لديهم بالفعل خلايا حساسة للضوء، ولكن لا توجد عدسة ولا وسيلة أخرى لإسقاط صورة على هذه الخلايا. يمكنهم التمييز بين النور والظلام، لكن ليس أكثر. هذا يمكّن القواقع من الابتعاد عن أشعة الشمس المباشرة. في الكائنات الحية التي تعيش بالقرب من فتحات أعماق البحار، تم تبسيط العيون المركبة بشكل ثانوي وتكييفها لرؤية الضوء الأحمر الذي تنتجه الفتحات الساخنة - وبهذه الطريقة يمكن للحوامل تجنب الغليان أحياء.[10]
أنواع
هناك عشرة تخطيطات مختلفة للعين - كل طريقة تكنولوجية لالتقاط صورة بصرية شائعة الاستخدام من قبل البشر، باستثناء عدسات الزوم وفرينل، تحدث في الطبيعة.[1] يمكن تصنيف أنواع العيون إلى "عيون بسيطة"، ذات سطح مقعر واحد مستقبِل للضوء، و "عيون مركبة"، والتي تشتمل على عدد من العدسات الفردية الموضوعة على سطح محدب.[1] لاحظ أن كلمة "بسيطة" لا تعني انخفاض مستوى التعقيد أو الحدة. في الواقع، يمكن تكييف أي نوع من العيون مع أي سلوك أو بيئة تقريبًا. القيود الوحيدة الخاصة بأنواع العين هي الدقة - تمنعها فيزياء العيون المركبة من تحقيق دقة أفضل من 1 درجة. أيضًا، يمكن أن تحقق عيون التراكب حساسية أكبر من العيون الموضعية، لذا فهي أكثر ملاءمة للمخلوقات التي تعيش في الظلام.[1] تنقسم العيون أيضًا إلى مجموعتين على أساس البناء الخلوي للمستقبلات الضوئية، حيث تكون الخلايا المستقبلة للضوء إما مستلقية (كما في الفقاريات) أو عيينة (عضو) . هاتان المجموعتان ليستا أحاديتين. تمتلك القراصات أيضًا خلايا مهدبة، [11] وبعض بطنيات الأقدام، [12] بالإضافة إلى بعض الحلقات تمتلك كليهما.[13]
تحتوي بعض الكائنات الحية على خلايا حساسة للضوء لا تفعل شيئًا سوى اكتشاف ما إذا كانت البيئة المحيطة بها فاتحة أم مظلمة، وهو ما يكفي لجذب إيقاعات الساعة البيولوجية . لا تعتبر هذه عيونًا لأنها تفتقر إلى البنية الكافية التي يمكن اعتبارها عضوًا، ولا تنتج صورة.[14]
هناك عشرة طرق مختلفة لمخرجات العين، في الواقع العين هي الوسيلة التي يتم بها التقاط الصورة الضوئية بالنسبة للإنسان. ويمكن تصنيف أنواع العين إلى "عيون بسيطة"، ذات سطح مقعر واحد مستقبل للضوء، و"عيون مركبة" وهي تضم عددا من العدسات محدبة فردية بحيث يتمكن الكائن الحي من الرؤيا بأكثر من زاوية. تتفاوت العيون من حيث البساطة والتعقيد حسب البيئة المحيطة.[15][16]
العين البدائية
وحدة معالجة الضوء الأساسية للعيون هي الخلية المستقبلة للضوء، وهي خلية متخصّصة تحتوي على نوعين من الجزيئات في الغشاء: أحدهما بروتين حساس للضوء يدعى الأوبسين، والآخر خضاب يميز الألوان يدعى حامل اللون. سُمِّيَت المجموعات المماثلة لهذه الخلايا "بقعًا عينيةً"، ويُعتَقد بأنّها تطورت بشكلٍ مستقلٍ في مكان ما بين 40 و 65 مرة. أتاحت البقع العينية هذه للحيوانات إدراكًا أوليًا جدًا للاتجاه ولشدة الضوء، ولكنها لم تكن كافيةً للتمييز بين الشيء وما يحيطه.
من الواضح أنّ تطوير نظام بصري يمكنه تمييز اتجاه الضوء ضمن بضعة درجات هو أمر أكثر صعوبة بكثير، إذ تمتلك مثل هذا النظام ست شعب فقط من بين ثلاث وثلاثين شعبة. ومع ذلك، تُشَكِّل هذه الشعب 96٪ من الأنواع الحية.
بدأت هذه النظم الضوئية المعقدة بالتزامن مع التقعّر التدريجي للرقع العينية متعددة الخلايا، مما أتاح القدرة على تمييز اتجاهات السطوع، الأمر الذي ازداد دقةً مع زيادة عمق التقعر. لم تكن الرقع العينية المسطحة فعّالة في تحديد اتجاه الضوء، لأنّ الشعاع الضوئي سينشط نفس الخلايا الحساسة للضوء بغض النظر عن اتجاهه، وعلى ذلك، سمح تقعر العيون المجوّفة بتمايز اتجاهي محدود بسبب اختلاف الخلايا التي سينشطها الضوء اعتمادًا على زاويته.
امتلكت الحلزونات القديمة العيون المجوّفة التي ظهرت في العصر الكامبري، ويمكن مشاهدتها اليوم لدى بعض الحلزونات وغيرها من اللافقاريات المعاصرة كالمستورقة. ومع ذلك، كانت هذه العين الأولية مفيدة في تحديد وجود أو غياب الضوء أكثر من تحديدها اتجاهه؛ تغيّر ذلك تدريجيًا مع تعمّق حفرة العين، وتزايد عدد الخلايا المستقبلة للضوء، الأمر الذي زوّد الكائن الحي بمعلوماتٍ بصريةٍ أدق بشكلٍ متزايد.
تسارع تطور العين خلال الانفجار الكامبري بشكل كبير، وشمل ذلك تطورًا جذريًا لكل من معالجة الصور وتحديد اتجاه الضوء.
أصبح دور تقليل عرض فتحة الضوء فيما بعد أكثر فعالية من زيادة عمق التقعر في تحسين الدقة البصرية، إذ وصلت الكائنات الحية عبر ذلك إلى التصوير الحقيقي، الأمر الذي سمح بإدراك دقيق للاتجاهات، إضافةً إلى إدراك قليل للأشكال. تتواجد هذه العيون حاليًا لدى النواتي، لكنها توفر دقة رديئة وتصويرًا خافتًا بسبب افتقارها للقرنية أو العدسة. مع ذلك، لا تزال تُشكّل تحسّنًا كبيرًا مقارنةّ بالعيون البدائية.
مَنَعَ نمو الخلايا الشفافة التلوث والإصابة بالطفيليات، الأمر الذي سمح لمحتويات الحجرة العينية بالتخصص ببطء ضمن خلط شفاف، وبالتالي امتلاك تحسينات كتصفية اللون، معامل الانكسار الأعلى، إعاقة الأشعة فوق البنفسجية، القدرة على العمل داخل وخارج الماء. قد تكون هذه الطبقة مرتبطة بالانسلاخ الذي تمر به بعض الطوائف الحيوانية، كما في حاملات المخالب مثلًا.
تكون القرنية -إضافةً للعدسات والخلطين الشفافين- مسؤولةً عن تجميع الضوء، والمساهمة في تركيزه على الجزء الخلفي لشبكية العين. تحمي القرنية مقلة العين وفي الوقت نفسه تكون مسؤولة عن نحو ثلثي إجمالي قوة الانكسار في العين.
أحد الأسباب الرئيسية المحتملة وراء قدرة العيون على التقاط مجموعة أطوال موجية ضيقة ومحددة من الطيف الكهرومغناطيسي -الطيف المرئي، هو تطوّر الحساسية الضوئية لدى أنواعٍ مائيةٍ في البدء؛ فقط الضوء الأخضر والأزرق المرئيين يمكنهما الانتقال عبر الماء. أثّرت خاصية تصفية الضوء هذه ذاتها على الحساسية الضوئية للنبات.
عيون غير مركبة
العيون البسيطة موجودة في كل مكان إلى حد ما، وقد تطورت العيون الحاملة للعدسات سبع مرات على الأقل في الفقاريات ورأسيات الأرجل والحلقيات والقشريات والمكعبات .[17][المصدر لا يؤكد ذلك]
عيون الحفرة
عيون الحفرة، والمعروفة أيضًا باسم العيون الجذعية، هي بقع للعين يمكن وضعها في حفرة لتقليل زوايا الضوء التي تدخل وتؤثر على بقعة العين، للسماح للكائن باستنتاج زاوية الضوء الوارد.[1] وجدت هذه الأشكال الأساسية في حوالي 85٪ من الشعب، وربما كانت مقدمة لأنواع أكثر تقدمًا من "العيون البسيطة". إنها صغيرة، وتتألف من حوالي 100 خلية تغطي حوالي 100 خلية µ م.[1] يمكن تحسين الاتجاه عن طريق تقليل حجم الفتحة، من خلال دمج طبقة عاكسة خلف خلايا المستقبل، أو عن طريق ملء الحفرة بمادة قابلة للكسر.[1]
طورت أفاعي الحفر حفرًا تعمل كعيون عن طريق استشعار الأشعة تحت الحمراء الحرارية، بالإضافة إلى عيونها ذات الطول الموجي البصري مثل عيون الفقاريات الأخرى (انظر استشعار الأشعة تحت الحمراء في الثعابين ). ومع ذلك، فإن أعضاء الحفرة مزودة بمستقبلات مختلفة نوعًا ما عن المستقبلات الضوئية، وهي قناة محتملة لمستقبلات عابرة محددة (قنوات TRP) تسمى TRPV1. الفرق الرئيسي هو أن المستقبلات الضوئية هي مستقبلات مقترنة ببروتين ج لكن TRP هي قنوات أيونية .
عين عدسة كروية
يمكن تحسين دقة عيون الحفرة بشكل كبير من خلال دمج مادة ذات معامل انكسار أعلى لتشكيل عدسة، مما قد يقلل بشكل كبير من نصف قطر التمويه الذي يتم مواجهته - وبالتالي زيادة الدقة التي يمكن الحصول عليها.[1] يتكون الشكل الأساسي، الذي يُرى في بعض بطنيات الأرجل والحلقيات، من عدسة ذات معامل انكسار واحد. يمكن الحصول على صورة أكثر وضوحًا باستخدام مواد ذات معامل انكسار عالٍ يتناقص حتى الحواف ؛ هذا يقلل من البعد البؤري وبالتالي يسمح لصورة حادة بالتشكل على الشبكية.[1] يسمح هذا أيضًا بفتحة أكبر للحصول على حدة معينة للصورة، مما يسمح بدخول المزيد من الضوء إلى العدسة ؛ وعدسة مسطحة، مما يقلل الزيغ الكروي .[1] هذه العدسة غير المتجانسة ضرورية حتى ينخفض البعد البؤري من حوالي 4 أضعاف نصف قطر العدسة إلى 2.5 نصف قطر.[1]
لقد تطورت العيون غير المتجانسة تسع مرات على الأقل: أربع مرات أو أكثر في بطنيات الأقدام، ومرة في مجدافيات الأرجل، ومرة في الحلقات، ومرة في رأسيات الأرجل، [1] ومرة في الكيتون، التي تحتوي على عدسات أراجونيت .[18] لا توجد كائنات مائية موجودة تمتلك عدسات متجانسة ؛ من المفترض أن يكون الضغط التطوري لعدسة غير متجانسة كبيرًا بما يكفي لكي "تتضخم" هذه المرحلة بسرعة.[1]
تخلق هذه العين صورة شديدة الوضوح لدرجة أن حركة العين يمكن أن تسبب ضبابية كبيرة. لتقليل تأثير حركة العين أثناء تحرك الحيوان، فإن معظم هذه العيون لها عضلات عين ثابتة.[1]
عين الحشرات تحمل عدسة بسيطة، لكن النقطة المحورية لها تقع دائمًا خلف شبكية العين ؛ وبالتالي، لا يمكنهم أبدًا تكوين صورة حادة. تعمل أوسيلي (عيون المفصليات من نوع الحفرة) على تعتيم الصورة عبر الشبكية بأكملها، وبالتالي فهي ممتازة في الاستجابة للتغيرات السريعة في شدة الضوء عبر المجال البصري بأكمله ؛ يتم تسريع هذه الاستجابة السريعة من خلال الحزم العصبية الكبيرة التي تندفع بالمعلومات إلى الدماغ.[19] قد يؤدي التركيز على الصورة أيضًا إلى تركيز صورة الشمس على عدد قليل من المستقبلات، مع احتمال حدوث ضرر تحت الضوء الشديد ؛ يحجب حجب المستقبلات بعض الضوء وبالتالي يقلل من حساسيتها.[19] أدت هذه الاستجابة السريعة إلى اقتراحات بأن عين الحشرات تستخدم بشكل أساسي أثناء الطيران، لأنه يمكن استخدامها للكشف عن التغيرات المفاجئة في الاتجاه الذي يرتفع فيه (لأن الضوء، وخاصة الضوء فوق البنفسجي الذي يمتصه الغطاء النباتي، يأتي عادةً من الأعلى) .[19]
عدسات متعددة
تحمل بعض الكائنات البحرية أكثر من عدسة. على سبيل المثال الكوبيبودا بونتيلا ثلاثة. السطح الخارجي له سطح مكافئ، يقاوم آثار الانحراف الكروي مع السماح بتشكيل صورة حادة. مجداف الأرجل الآخر، كوبيليا، له عدستان في كل عين، مرتبة مثل تلك الموجودة في التلسكوب.[1] مثل هذه الترتيبات نادرة وغير مفهومة جيدًا، ولكنها تمثل بناءًا بديلاً.
تُرى عدسات متعددة في بعض الصيادين مثل النسور والعناكب القافزة، التي لديها قرنية انكسارية: هذه لها عدسة سلبية، مما يوسع الصورة المرصودة بنسبة تصل إلى 50٪ فوق الخلايا المستقبلة، وبالتالي يزيد من الدقة البصرية.[1]
القرنية الانكسارية
في عيون معظم الثدييات والطيور والزواحف ومعظم الفقاريات الأرضية الأخرى (مع العناكب وبعض يرقات الحشرات) يكون للسائل الزجاجي معامل انكسار أعلى من الهواء.[1] بشكل عام، العدسة ليست كروية. عدسات كروية تنتج انحراف كروية. في القرنيات الانكسارية، يتم تصحيح أنسجة العدسة باستخدام مادة عدسة غير متجانسة (انظر عدسة لونبورغ )، أو بشكل شبه كروي.[1] تسطيح العدسة له عيب ؛ تتضاءل جودة الرؤية بعيدًا عن محور التركيز الرئيسي. وبالتالي، فإن الحيوانات التي تطورت بمجال رؤية واسع غالبًا ما يكون لها عيون تستخدم عدسة غير متجانسة.[1]
كما ذكر أعلاه، القرنية الانكسارية مفيدة فقط خارج الماء. في الماء، هناك اختلاف بسيط في معامل الانكسار بين السائل الزجاجي والماء المحيط. ومن ثم، فإن الكائنات التي عادت إلى الماء - طيور البطريق والفقمات، على سبيل المثال - تفقد قرنيتها شديدة التقوس وتعود إلى الرؤية القائمة على العدسة. الحل البديل، الذي يتحمله بعض الغواصين، هو أن تكون القرنية شديدة التركيز.[1]
عيون عاكسة
من البدائل للعدسة محاذاة داخل العين بـ "مرايا"، وتعكس الصورة للتركيز عند نقطة مركزية.[1] إن طبيعة هذه العيون تعني أنه إذا نظر المرء إلى بؤبؤ عين، سيرى نفس الصورة التي يراها الكائن الحي، تنعكس مرة أخرى.[1]
تستخدم العديد من الكائنات الحية الصغيرة مثل الروتيفر ومجدافيات الأرجل والديدان المفلطحة مثل هذه الأعضاء، ولكنها أصغر من أن تنتج صورًا قابلة للاستخدام.[1] بعض الكائنات الحية الكبيرة، مثل الأسقلوب، تستخدم أيضًا عيونًا عاكسة. تحتوي المحارات الصدفية بكتين (ذوات الصدفتين) على عيون عاكسة بمقياس 100 ملم تهدد حافة غلافه. يكتشف الأجسام المتحركة أثناء مرورها بعدسات متتالية.[1]
يوجد على الأقل حيوان فقاري واحد، وهو السمكة الشبحية، والتي تشتمل عيونها على بصريات عاكسة لتركيز الضوء. تجمع كل من عينا سمكة شبح الضوء من الأعلى والأسفل ؛ يتم تركيز الضوء القادم من الأعلى بواسطة عدسة، بينما يأتي الضوء من الأسفل بواسطة مرآة منحنية تتكون من عدة طبقات من الألواح العاكسة الصغيرة المصنوعة من بلورات الجوانين .[20]
عيون مركبة



قد تتكون العين المركبة من آلاف وحدات مستقبلات الضوء الفردية أو أوماتيديوم ( أوماتيديوم، المفرد). الصورة المدركة هي مزيج من المدخلات من العديد من وماتيديوم( "وحدات العين" الفردية)، والتي تقع على سطح محدب، وبالتالي تشير في اتجاهات مختلفة قليلاً. بالمقارنة مع العيون البسيطة، تمتلك العيون المركبة زاوية رؤية كبيرة جدًا، ويمكنها اكتشاف الحركة السريعة، وفي بعض الحالات، استقطاب الضوء.[21] نظرًا لأن العدسات الفردية صغيرة جدًا، فإن تأثيرات الانعراج تفرض حدًا على الدقة المحتملة التي يمكن الحصول عليها (بافتراض أنها لا تعمل كمصفوفات مرحلية ). لا يمكن مواجهة ذلك إلا من خلال زيادة حجم العدسة وعددها. لنرى بدقة مماثلة لأعيننا البسيطة، سيحتاج البشر إلى عيون مركبة كبيرة جدًا، حوالي 11 متر (36 قدم) في دائرة نصف قطرها.[22]
تنقسم العيون المركبة إلى مجموعتين: العيون الموضعية، والتي تشكل صورًا مقلوبة متعددة، وعيون التراكب، والتي تشكل صورة منتصبة واحدة.[23] العيون المركبة شائعة في المفصليات، والحلقيات وبعض الرخويات ذات الصدفتين.[24] تنمو العيون المركبة في مفصليات الأرجل على هوامشها بإضافة أوماتيديوم جديدة.[25]
عيون الموقف

عيون المظهر هي الشكل الأكثر شيوعًا للعيون ويفترض أنها الشكل السلفي للعيون المركبة. توجد في جميع مجموعات المفصليات، على الرغم من أنها قد تطورت أكثر من مرة داخل هذه الشعبة.[1] تحتوي بعض الحلقات وذوات الصدفتين أيضًا على عيون. يمتلكها أيضًا ليمولوس، سلطعون حدوة الحصان، وهناك اقتراحات بأن المخلّبات الآخرين طوروا عيونهم البسيطة عن طريق الاختزال من نقطة البداية المركبة.[1] (يبدو أن بعض اليرقات قد طورت عيونًا مركبة من عيون بسيطة في الاتجاه المعاكس. )
تعمل عيون المظهر من خلال جمع عدد من الصور، واحدة من كل عين، ودمجها في الدماغ، حيث تساهم كل عين عادة بنقطة واحدة من المعلومات. وبدل العين النموذجية لديها عدسة التركيز ضوء من اتجاه واحد على رابدوم، في حين يتم امتصاص الضوء من اتجاهات أخرى من الجدار الظلام من مقيلة .
عيون التراكب
يسمى النوع الثاني تراكب العين. تنقسم عين التراكب إلى ثلاثة أنواع:
- الانكسار.
- يعكس.
- تراكب مكافئ.
العين المتراكبة الانكسارية بها فجوة بين العدسة والهيكل، ولا يوجد جدار جانبي. تأخذ كل عدسة الضوء بزاوية على محورها وتعكسه بنفس الزاوية على الجانب الآخر. والنتيجة هي صورة نصف قطرها نصف قطر العين، حيث توجد أطراف الممرات. هذا النوع من العين المركبة، الذي يوجد له حجم ضئيل لا يمكن أن يحدث تراكب فعال تحته، [26] يوجد عادة في الحشرات الليلية، لأنه يمكن أن يخلق صورًا أكثر سطوعًا 1000 مرة من عيون الموضع المكافئة، على الرغم من تكلفة تقليل الدقة.[27] في نوع العين المركب ذو التراكب المكافئ، الذي يُرى في المفصليات مثل الذباب، تركز الأسطح المكافئة لداخل كل وجه من الضوء من عاكس إلى مجموعة أجهزة استشعار. منذ فترة طويلة جسديا القشريات العشارى الأرجل مثل الروبيان، الجمبري، جراد البحر والكركند هي وحدها في وجود يعكس عيون تراكب، والتي لها أيضا وجود فجوة ولكن استخدام الزاوية شفافة المرايا بدلا من العدسات.
تراكب مكافئ
يعمل نوع العين هذا عن طريق انكسار الضوء، ثم استخدام مرآة مكافئة لتركيز الصورة ؛ فهو يجمع بين ميزات التراكب والعيون الموضعية.[10]
آخرى
نوع آخر من العيون المركبة، يوجد في الذكور من رتبة ملتويات الأجنحة، يستخدم سلسلة من العيون البسيطة - عيون لها فتحة واحدة توفر الضوء لشبكية العين التي تشكل صورة كاملة. تشكل العديد من هذه الثقوب معًا عين مركب ستريبسيبتيران، والتي تشبه العيون المركبة `` الفصامية '' لبعض ثلاثية الفصوص .[28] لأن كل ثقب هو عين بسيطة، فإنه ينتج صورة مقلوبة ؛ يتم دمج هذه الصور في الدماغ لتشكيل صورة واحدة موحدة. نظرًا لأن فتحة العيينة أكبر من جوانب العين المركبة، فإن هذا الترتيب يسمح بالرؤية في ظل مستويات الإضاءة المنخفضة.[1]
تحتوي المنشورات الجيدة مثل الذباب أو نحل العسل، أو الحشرات التي تصطاد الفرائس مثل فرس النبي أو اليعسوب، على مناطق متخصصة من الأوماتيديا منظمة في منطقة نقرة تعطي رؤية حادة. في المنطقة الحادة، العيون مفلطحة والأوجه أكبر. يسمح التسطيح لمزيد من الأوماتيديا باستقبال الضوء من بقعة وبالتالي دقة أعلى. تسمى البقعة السوداء التي يمكن رؤيتها على العيون المركبة لمثل هذه الحشرات، والتي تبدو دائمًا وكأنها تنظر مباشرة إلى المراقب، بالبؤبؤ الكاذب . يحدث هذا لأن الأوماتيديا التي يلاحظها المرء "وجهاً لوجه" (على طول محاورها البصرية ) تمتص الضوء الساقط، بينما تعكسه تلك الموجودة على جانب واحد.[29]
وهناك بعض الاستثناءات من الأنواع المذكورة أعلاه. تحتوي بعض الحشرات على ما يسمى بالعين المركبة ذات العدسة المفردة، وهي نوع انتقالي بين نوع تراكب للعين المركبة متعددة العدسات والعين ذات العدسة المفردة الموجودة في الحيوانات ذات العيون البسيطة. ثم هناك مايسيد الروبيان، انحلال البول السكري. لدى الجمبري عين من نوع التراكب الانكساري، في المؤخرة خلف كل عين يوجد وجه واحد كبير يبلغ قطره ثلاثة أضعاف قطر الآخر في العين وخلفه يوجد مخروط بلوري متضخم. هذا يعرض صورة منتصبة على شبكية العين المتخصصة. العين الناتجة هي مزيج من عين بسيطة داخل عين مركبة.
نسخة أخرى هي عين مركبة يشار إليها غالبًا باسم "سيودوفاستيد"، كما رأينا في سكوتيجيرا.[30] يتكون هذا النوع من العين من مجموعة من العديد من الأوماتيديا على كل جانب من الرأس، منظمة بطريقة تشبه العين المركبة الحقيقية.
جسد أوبوفاياكومو وينتي، وهو نوع من النجوم الهشة، مغطى بـ أوماتيديا، مما يحول جلده بالكامل إلى عين مركبة. نفس الشيء ينطبق على العديد من الكيتون . تحتوي الأقدام الأنبوبية لقنافذ البحر على بروتينات مستقبِلة للضوء تعمل معًا كعين مركبة ؛ تفتقر إلى أصباغ الفرز، لكن يمكنها اكتشاف اتجاه الضوء بواسطة الظل الذي يلقي به جسمها المعتم.[31]
العناصر الغذائية
الجسم الهدبي مثلث في المقطع الأفقي ومغطى بطبقة مزدوجة، الظهارة الهدبية. الطبقة الداخلية شفافة وتغطي الجسم الزجاجي، ومستمرة من النسيج العصبي لشبكية العين. الطبقة الخارجية شديدة التصبغ، متصلة بالظهارة الصبغية للشبكية، وتشكل خلايا العضلة الموسعة.
الجسم الزجاجي هو كتلة هلامية شفافة عديمة اللون تملأ الفراغ بين عدسة العين وشبكية العين التي تبطن مؤخرة العين.[32] انا تنتج بواسطة خلايا معينة. إنه ذو تركيب مشابه إلى حد ما للقرنية، ولكنه يحتوي على عدد قليل جدًا من الخلايا (معظمها خلايا بلعمية تزيل الحطام الخلوي غير المرغوب فيه في المجال البصري، بالإضافة إلى الخلايا الهيالوسية لبالاز على سطح الجسم الزجاجي، والتي تعيد معالجة حمض الهيالورونيك) الأوعية الدموية، و 98-99٪ من حجمها عبارة عن ماء (مقابل 75٪ في القرنية) مع أملاح، سكريات، فيتروسين (نوع من الكولاجين)، شبكة من ألياف الكولاجين من النوع الثاني مع حمض الهيالورونيك عديد السكاريد المخاطي، و أيضا مجموعة واسعة من البروتينات بكميات صغيرة. بشكل مثير للدهشة، مع وجود القليل من المادة الصلبة، فإنه يمسك العين.
تشريح عين الإنسان
 مقالة مفصلة: عين الإنسان
مقالة مفصلة: عين الإنسان
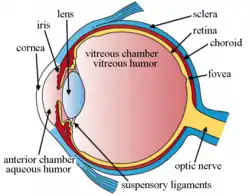
تُقسم عين الإنسان تشريحياً إلى ثلاث طبقات بالإضافة لأوساط كاسرة للضوء:[33]
- طبقة خارجية ليفية: القرنية + الصلبة.
- طبقة متوسطة وعائية صباغية: جسم هدبي + مشيمية + قزحية.
- طبقة داخلية عصبية: شبكية.
الطبقة الليفية الخارجية:
تشكل الصلبة القسم غير الشفاف منها، وتُكوِّن 85% من الطبقة الخارجية، في حين تشكل القرنية القسم الشفاف وتُكوِّن 15% من الطبقة الخارجية.
الطبقة المتوسطة الوعائية الصباغية:
تتألف من المشيمية وهي غشاء قاتم يرتبط بشكل متين بالشبكية، والجسم الهدبي وهو عبارة عن بنية عضلية وعائية تساهم في عملية المطابقة بالإضافة لزوائد هدبية تفرز سائل مائي القوام وألياف متينة ترتبط مع العدسة، أما القزحية فهي بنية صباغية تحتوي عضلات مُقبضة ومُوسعة يتغير حجمها بحسب كمية الضوء التي تتعرض لها العين، وتقسم القزحية العين إلى حجرتين أمامية وخلفية.
الطبقة الداخلية العصبية:
تتألف من الشبكية التي تُقسم بدورها إلى: قسم بصري (عصي ومخاريط) وقسم غير بصري هدبي قزحي، وتعد الشبكية استمراراً للعصب البصري وفيها طبقتان إحداهما عصبية والأخرى صباغية وبينهما فراغ يُسمى الحيز داخل الشبكية.
تطور
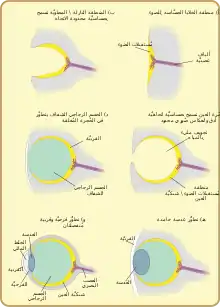
يعتبر الاستقبال الضوئي قديمًا جدًا من الناحية التطورية، مع نظريات مختلفة عن التطور.[34] أصبح الأصل المشترك ( الأحادي ) لجميع عيون الحيوانات مقبولًا على نطاق واسع كحقيقة. هذا يعتمد على السمات الجينية المشتركة لجميع العيون ؛ أي أن جميع العيون الحديثة، متنوعة كما هي، لها أصولها في عين بدائية يُعتقد أنها تطورت منذ حوالي 540 مليون سنة، [35][36][37] ويعتبر جين باكس 6 عاملاً رئيسياً في هذا . يُعتقد أن غالبية التطورات في العيون المبكرة قد استغرق تطويرها بضعة ملايين من السنين فقط، حيث كان أول حيوان مفترس يحصل على تصوير حقيقي قد أطلق "سباق تسلح" [38] بين جميع الأنواع التي لم تهرب من الصورة الضوئية بيئة. ستكون الحيوانات الفريسة والحيوانات المفترسة المتنافسة على حد سواء في وضع غير موات بشكل واضح بدون هذه القدرات وستكون أقل احتمالا للبقاء والتكاثر. ومن ثم تطورت أنواع وأنواع فرعية متعددة من العين بالتوازي (باستثناء تلك الخاصة بالمجموعات، مثل الفقاريات، التي تم إجبارها فقط على البيئة الضوئية في مرحلة متأخرة)
تظهر العيون في الحيوانات المختلفة التكيف مع متطلباتها. على سبيل المثال، تتمتع عين طائر جارح بحدة بصرية أكبر بكثير من عين الإنسان، وفي بعض الحالات يمكنها اكتشاف الأشعة فوق البنفسجية. تعتبر الأشكال المختلفة للعين، على سبيل المثال، الفقاريات والرخويات أمثلة على التطور المتوازي، على الرغم من أصلها المشترك البعيد. التقارب المظهري لهندسة رأسيات الأرجل ومعظم عيون الفقاريات يخلق انطباعًا بأن عين الفقاريات قد تطورت من تصوير عين رأسي الأرجل، ولكن هذا ليس هو الحال، حيث إن الأدوار المعكوسة لفئتي opsin الهدبية والرباطية الخاصة بها [39] وعدسة مختلفة تظهر البلورات.[40]
كانت أقدم "العيون"، والتي تسمى بقع العين، عبارة عن بقع بسيطة من البروتين المستقبِل للضوء في الحيوانات وحيدة الخلية. في الكائنات متعددة الخلايا، تطورت بقع عينية متعددة الخلايا، تشبه فيزيائيًا بقع مستقبلات الذوق والشم. يمكن لبقع العين هذه أن تشعر فقط بالسطوع المحيط: يمكنها التمييز بين الضوء والظلام، ولكن ليس اتجاه مصدر الضوء.[1]
من خلال التغيير التدريجي، تنخفض نقاط أعين الأنواع التي تعيش في بيئات مضاءة جيدًا إلى شكل "كوب" ضحل. تم تحقيق القدرة على التمييز بشكل طفيف في السطوع الاتجاهي باستخدام الزاوية التي يصطدم فيها الضوء بخلايا معينة لتحديد المصدر. تعمقت الحفرة بمرور الوقت، وتضاءلت الفتحة في الحجم، وزاد عدد الخلايا المستقبلة للضوء، مما شكل كاميرا ذات ثقب فعال قادرة على تمييز الأشكال بشكل باهت.[41] ومع ذلك، فإن أسلاف الحديث سمك الجريث، ويعتقد أن يكون الفقاريات، [39] وقد دفعت الواضح أن عميقة جدا، والمياه الظلام، حيث كانوا أقل عرضة للالضواري النظر، وحيث أنه من المفيد أن يكون محدب العين بقعة، التي يجمع ضوءًا أكثر من الضوء المسطح أو المقعر. كان من الممكن أن يؤدي هذا إلى مسار تطوري مختلف إلى حد ما لعين الفقاريات عن عيون الحيوانات الأخرى.
النمو الزائد الرقيق للخلايا الشفافة فوق فتحة العين، والذي تم تشكيله في الأصل لمنع تلف بقعة العين، سمح للمحتويات المنفصلة في حجرة العين بالتخصص في روح الدعابة الشفافة التي تعمل على تحسين ترشيح الألوان، ومنع الإشعاع الضار، وتحسين معامل الانكسار للعين، و يسمح بالوظيفة خارج الماء. تنقسم الخلايا الواقية الشفافة في النهاية إلى طبقتين، مع وجود سائل في الدورة الدموية بينهما مما سمح بزوايا رؤية أوسع ودقة تصوير أكبر، وزاد سمك الطبقة الشفافة تدريجياً، في معظم الأنواع التي تحتوي على بروتين بلوري شفاف.[42]
شكلت الفجوة بين طبقات الأنسجة بشكل طبيعي شكلًا محدبًا، وهو هيكل مثالي على النحو الأمثل لمؤشر انكسار عادي بشكل مستقل، تنقسم طبقة شفافة وطبقة غير شفافة إلى الأمام من العدسة: القرنية والقزحية . أدى فصل الطبقة الأمامية مرة أخرى إلى تكوين روح الدعابة، الخلط المائي . هذه الزيادة في القوة الانكسارية، ومرة أخرى خففت مشاكل في الدورة الدموية. يسمح تكوين حلقة غير شفافة لمزيد من الأوعية الدموية، ومزيد من الدورة الدموية، وحجم أكبر للعين.[42]
العلاقة بمتطلبات الحياة
تتكيف العيون بشكل عام مع البيئة ومتطلبات الحياة للكائن الحي الذي يحملها. على سبيل المثال، يميل توزيع المستقبلات الضوئية إلى مطابقة المنطقة التي تتطلب أعلى حدة، مع الكائنات الحية التي تقوم بمسح الأفق، مثل تلك التي تعيش في السهول الأفريقية، والتي لها خط أفقي من العقد عالية الكثافة، بينما يسكن الأشجار. تميل المخلوقات التي تتطلب رؤية شاملة جيدة إلى أن يكون لها توزيع متماثل للعقد، مع انخفاض حدة الحدة للخارج من المركز.
بطبيعة الحال، بالنسبة لمعظم أنواع العيون، من المستحيل الابتعاد عن الشكل الكروي، لذلك يمكن فقط تغيير كثافة المستقبلات الضوئية. في الكائنات الحية ذات العيون المركبة، فإن عدد الأوماتيديا بدلاً من العقد هو الذي يعكس منطقة الحصول على أعلى البيانات.[1] :23–24 إن عيون التراكب البصري مقيدة بشكل كروي، ولكن الأشكال الأخرى من العيون المركبة قد تتشوه إلى شكل حيث يتم محاذاة المزيد من الأوماتيديا، على سبيل المثال، الأفق، دون تغيير حجم أو كثافة الأوماتيديا الفردية.[43] عيون الكائنات التي تقوم بمسح الأفق لها سيقان بحيث يمكن محاذاة بسهولة مع الأفق عندما يكون هذا مائلاً، على سبيل المثال، إذا كان الحيوان على منحدر.[29]
امتداد لهذا المفهوم هو أن عيون الحيوانات المفترسة عادة ما يكون لها منطقة رؤية حادة للغاية في مركزها، للمساعدة في تحديد الفريسة.[43] في كائنات المياه العميقة، قد لا يكون مركز العين متضخمًا. و هابيرياويات مزدوجات الأرجل حيوانات المياه العميقة التي تتغذى على الكائنات الحية فوقهم. تنقسم عيونهم تقريبًا إلى قسمين، ويُعتقد أن المنطقة العلوية تشارك في الكشف عن الصور الظلية للفريسة المحتملة - أو الحيوانات المفترسة - مقابل الضوء الخافت للسماء أعلاه. وبناءً على ذلك، فإن هيبرييدات المياه العميقة، حيث يجب مقارنة الضوء الذي يجب مقارنة الصور الظلية به يكون باهتًا، ولها "عيون علوية" أكبر، وقد تفقد الجزء السفلي من أعينها تمامًا.[43] في القطب الجنوبي العملاق جليبتونوتوس أنتاركتيكوس، يتم فصل العين المركبة البطنية الصغيرة تمامًا عن العين المركبة الظهرية الأكبر بكثير.[44] يمكن تحسين إدراك العمق من خلال تكبير العيون في اتجاه واحد ؛ تشويه العين قليلاً يسمح بتقدير المسافة إلى الجسم بدرجة عالية من الدقة.[10]
تكون البراعة أعلى بين الكائنات الحية الذكرية التي تتزاوج في الهواء، حيث يجب أن تكون قادرة على تحديد وتقييم الزملاء المحتملين على خلفية كبيرة جدًا.[43] من ناحية أخرى، فإن عيون الكائنات الحية التي تعمل في مستويات الإضاءة المنخفضة، مثل وقت الفجر والغسق أو في المياه العميقة، تميل إلى أن تكون أكبر لزيادة كمية الضوء التي يمكن التقاطها.[43]
ليس شكل العين فقط هو الذي قد يتأثر بنمط الحياة. يمكن أن تكون العيون أكثر الأجزاء المرئية من الكائنات الحية، وهذا يمكن أن يكون بمثابة ضغط على الكائنات الحية للحصول على عيون أكثر شفافية على حساب الوظيفة.[43]
يمكن تثبيت العيون على سيقان لتوفير رؤية شاملة أفضل، عن طريق رفعها فوق درع الكائن الحي ؛ هذا يسمح لهم أيضًا بتتبع الحيوانات المفترسة أو الفريسة دون تحريك الرأس.[10]
علم وظائف الأعضاء
حدة البصر

حدة البصر، أو القدرة على التحليل، هي "القدرة على التمييز بين التفاصيل الدقيقة" وهي خاصية للخلايا المخروطية .[32] غالبًا ما يتم قياسه في دورات لكل درجة (CPD)، والتي تقيس الدقة الزاوية، أو مدى قدرة العين على تمييز كائن عن آخر من حيث الزوايا المرئية. يمكن قياس الدقة في CPD من خلال المخططات الشريطية لأعداد مختلفة من دورات الشريط الأبيض / الأسود. على سبيل المثال، إذا كان كل نمط 1.75 بعرض سم ويوضع على مسافة 1 متر من العين، فإنه يقابل زاوية مقدارها 1 درجة، وبالتالي فإن عدد أزواج الأعمدة البيضاء / السوداء على النموذج سيكون مقياسًا للدورات لكل درجة من هذا النمط. أعلى رقم يمكن للعين حله على شكل خطوط أو تمييزه عن الكتلة الرمادية هو قياس حدة البصر للعين.
بالنسبة للعين البشرية ذات حدة ممتازة، فإن الحد الأقصى للدقة النظرية هو 50 CPD [45] (1.2 دقيقة قوسية لكل زوج خط، أو 0.35 زوج خط مم، عند 1 م). يمكن للفأر أن يحل فقط حوالي 1 إلى 2 CPD.[46] يمتلك الحصان حدة أعلى من خلال معظم المجال البصري لعينيه مقارنة بالإنسان، ولكنه لا يتناسب مع حدة منطقة النقرة المركزية للعين البشرية.[47]
الانحراف الكروي يحد من دقة 7 ملم تلميذ إلى حوالي 3 قوسين لكل زوج خط. عند قطر التلميذ 3 مم، يتم تقليل الانحراف الكروي بشكل كبير، مما ينتج عنه دقة محسنة تقارب 1.7 دقيقة قوسية لكل زوج خط.[48] دقة 2 قوس لكل زوج خط، أي ما يعادل فجوة 1 قوس دقيقة في نمط بصري، يتوافق مع 20/20 (رؤية طبيعية) في البشر
ومع ذلك، في العين المركبة، يرتبط الدقة بحجم الأوماتيديا الفردية والمسافة بين الأوماتيديا المجاورة. جسديًا لا يمكن تصغير حجمها لتحقيق حدة الرؤية بالعين ذات العدسة الواحدة كما في الثدييات. العيون المركبة لديها حدة أقل بكثير من عيون الفقاريات.[49]
إدراك اللون
"رؤية اللون هي كلية الكائن الحي لتمييز الأضواء ذات الصفات الطيفية المختلفة." [32] تقتصر جميع الكائنات على نطاق صغير من الطيف الكهرومغناطيسي ؛ هذا يختلف من مخلوق إلى مخلوق، ولكنه يتراوح بين أطوال موجية 400 و 700 نانومتر.[50] هذا جزء صغير نوعًا ما من الطيف الكهرومغناطيسي، ربما يعكس تطور الغواصة للعضو: يحجب الماء جميع النوافذ الصغيرة باستثناء نافذتين صغيرتين من الطيف الكهرومغناطيسي، ولم يكن هناك ضغط تطوري بين الحيوانات البرية لتوسيع هذا النطاق.[51]
أكثر الأصباغ حساسية، رودوبسين، لها ذروة استجابة عند 500 نانومتر.[52] يمكن للتغييرات الصغيرة في ترميز الجينات لهذا البروتين تعديل ذروة الاستجابة ببضعة نانومتر ؛ [2] للأصباغ الموجودة في العدسة أيضًا تصفية الضوء الوارد، وتغيير ذروة الاستجابة.[2] العديد من الكائنات الحية غير قادرة على التمييز بين الألوان، وبدلاً من ذلك ترى في ظلال رمادية ؛ تتطلب رؤية الألوان مجموعة من الخلايا الصبغية التي تكون حساسة بشكل أساسي لنطاقات أصغر من الطيف. في الرئيسيات والأبراص والكائنات الحية الأخرى، تأخذ هذه شكل خلايا مخروطية، تطورت منها الخلايا العصوية الأكثر حساسية.[52] حتى لو كانت الكائنات الحية قادرة جسديًا على التمييز بين الألوان المختلفة، فإن هذا لا يعني بالضرورة أنها تستطيع إدراك الألوان المختلفة ؛ فقط مع الاختبارات السلوكية يمكن استنتاج ذلك.[2]
يمكن لمعظم الكائنات الحية ذات الرؤية اللونية الكشف عن الأشعة فوق البنفسجية. هذا الضوء ذو الطاقة العالية يمكن أن يضر بالخلايا المستقبلة. مع استثناءات قليلة (الثعابين والثدييات المشيمية)، تتجنب معظم الكائنات الحية هذه الآثار من خلال وجود قطرات زيت ماصة حول خلاياها المخروطية. البديل، الذي طورته الكائنات الحية التي فقدت قطرات الزيت هذه أثناء التطور، هو جعل العدسة منيعة أمام الضوء فوق البنفسجي - وهذا يحول دون إمكانية اكتشاف أي ضوء فوق البنفسجي، لأنه لا يصل حتى إلى شبكية العين.[52]
قضبان ومخاريط
تحتوي شبكية العين على نوعين رئيسيين من خلايا المستقبلات الضوئية الحساسة للضوء والمستخدمة للرؤية: العصي والمخاريط.
لا يمكن للقضبان تمييز الألوان، ولكنها مسؤولة عن الرؤية أحادية اللون في الإضاءة المنخفضة ( الاسكتلندية ) ( أبيض وأسود ) أنها تعمل بشكل جيد في ضوء خافت لأنها تحتوي على صبغة، رودوبسين التشبع (البصرية اللون الأرجواني)، وهي حساسة في شدة الإضاءة الخافتة، ولكن على ارتفاع ( ضيائية ) شدة. يتم توزيع العصي في جميع أنحاء الشبكية ولكن لا يوجد أي منها عند النقرة ولا يوجد أي منها في النقطة العمياء . كثافة القضيب أكبر في شبكية العين المحيطية منها في شبكية العين المركزية.
المخاريط هي المسؤولة عن رؤية الألوان . فهي تتطلب ضوء أكثر إشراقا للعمل مما تقتضيه قضبان. في البشر، هناك ثلاثة أنواع من المخاريط، وهي حساسة إلى أقصى حد لطول الموجة الطويلة، وطول الموجة المتوسطة، والضوء قصير الموجة (يشار إليها غالبًا بالأحمر والأخضر والأزرق على التوالي، على الرغم من أن قمم الحساسية ليست في الواقع عند هذه الألوان ). اللون المرئي هو التأثير المشترك للمنبهات والاستجابات من هذه الأنواع الثلاثة من الخلايا المخروطية. تتركز غالب المخاريط في الاقماع وقرب النقرة. يوجد عدد قليل فقط على جانبي الشبكية. تُرى الأشياء بشكل أكثر حدة في التركيز البؤري عندما تقع صورها على النقرة، كما هو الحال عندما ينظر المرء إلى كائن ما مباشرة. ترتبط الخلايا والقضبان المخروطية من خلال الخلايا الوسيطة في شبكية العين بالألياف العصبية للعصب البصري . عندما يتم تحفيز العصي والمخاريط بالضوء، فإنها تتصل عبر الخلايا المجاورة داخل الشبكية لإرسال إشارة كهربائية إلى ألياف العصب البصري. ترسل الأعصاب البصرية نبضات عبر هذه الألياف إلى الدماغ.[52]
تصبغ
تتنوع جزيئات الصبغة المستخدمة في العين، ولكن يمكن استخدامها لتحديد المسافة التطورية بين المجموعات المختلفة، ويمكن أيضًا أن تكون مساعدة في تحديد أيها وثيق الصلة - على الرغم من وجود مشاكل التقارب.[52]
أوبسينس هي الأصباغ المشاركة في استقبال الضوء. تستخدم أصباغ أخرى، مثل الميلانين، لحماية الخلايا المستقبلة للضوء من الضوء المتسرب من الجانبين. تطورت مجموعة بروتين أوبسينس قبل فترة طويلة من آخر سلف مشترك للحيوانات، واستمرت في التنويع منذ ذلك الحين.[2]
هناك نوعان من أوبسينس تشارك في الرؤية ؛ سي-أوبسينس، التي ترتبط بخلايا مستقبلة للضوء من النوع الهدبي، و آر-أوبسينس، المرتبطة بخلايا مستقبلات الضوء رابدوميرك (ربومي).[53] عادة ما تحتوي عيون الفقاريات على خلايا هدبية مع سي-أوبسينس، و (ثنائي) اللافقاريات لها خلايا رابدوميرك في العين مع آر-أوبسينس. ومع ذلك، فإن بعض الخلايا العقدية للفقاريات تعبر عن آر-أوبسينس، مما يشير إلى أن أسلافهم استخدموا هذا الصباغ في الرؤية، وأن البقايا تبقى حية في العين.[53] وبالمثل، تم العثور على سي-أوبسينس ليتم التعبير عنها في دماغ بعض اللافقاريات. ربما تم التعبير عنها في الخلايا الهدبية لعيون اليرقات، والتي تم إعادة امتصاصها لاحقًا في الدماغ عند التحول إلى شكل البالغين.[53] تم العثور على سي-أوبسينس أيضًا في بعض العيون المشتقة من اللافقاريات الثنائية، مثل العيون الشاحبة للرخويات ذات الصدفتين ؛ ومع ذلك، فإن العيون الجانبية (التي من المفترض أنها من النوع السلفي لهذه المجموعة، إذا تطورت العيون مرة واحدة هناك) تستخدم دائمًا آر-أوبسينس.[53] لاسعات، وهي مجموعة خارجية إلى الأصناف المذكورة أعلاه، صريحة سي-أوبسينس - ولكن آر-أوبسينس لم يتم العثور عليها بعد في هذه المجموعة.[53] بالمناسبة، يتم إنتاج الميلانين المنتج في القنيات بنفس الطريقة التي يتم إنتاجها في الفقاريات، مما يشير إلى الأصل المشترك لهذا الصباغ.[53]
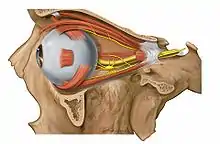
الأوساط الكاسرة للضوء
تحوي العين مجموعة من الأوساط الكاسرة للضوء تساهم في حدوث الرؤية بشكل صحيح [54] وهذه الأوساط هي:
- القرنية: بنية شفافة لا تحتوي أوعية دموية مسؤولة عن انكسار الضوء.
- الجسم المائي: تفرزه الزوائد الهدبية ويتألف من حجرتين أمامية وخلفية تفصل بينهما القزحية.
- العدسة: بنية شفافة محدبة تتواجد داخل محفظة ليفية ومعلقة بالرباط المعلق. المهمة الرئيسية للعدسة تركيز صورة الأجسام المرئية في بؤرة العين، فإذا كان الجسم الهدبي مسترخياً سيؤدي ذلك لتوتير الأربطة وتسطيح العدسة، أما إذا كان الجسم الهدبي متقلصاً سيؤدي ذلك لارتخاء الأربطة وتكوير العدسة.
- الجسم الزجاجي: وسط شفاف شبيه بالهلام يُشكل بمفرده 80% من كرة العين، مهمته ضبط الشبكية ودعم العدسة.
أعصاب العين وأوعيتها
للعين عصب حسي واحد هو العصب البصري، وثلاثة أعصاب حركية هي العصب المحرك للعين والعصب البكري والعصب المُبعد.[33][54]
- العصب البصري: العصب القحفي الثاني يمر عبر النفق البصري وينتهي بالشبكية تغلفه السحايا ويعتبر عصباً حسياً مسؤولاً عن حاسة الرؤية.
- العصب المحرك للعين: العصب القحفي الثالث يمر من الشق الحجاجي العلوي وله فرعان علوي وسفلي. يعتبر عصباً حركياً مسؤولاً عن تعصيب جميع عضلات العين عدا العضلة المستقيمة الوحشية والعضلة المنحرفة العلوية، بالإضافة لألياف نظير ودية تعصب العضلة المُقبضة للعين والعضلة الهدبية.
- العصب البكري: العصب القحفي الرابع. ينشأ من الوجه الخلفي لجذع الدماغ ويمر عبر الشق الحجاجي العلوي ويعصب العضلة المنحرفة العلوية.
- العصب المُبعد: العصب القحفي السادس. ينشأ من منتصف جذع الدماغ ويمر عبر الشق الحجاجي العلوي ويعصب العضلة المستقيمة الوحشية.
تأتي التروية الدموية للعين من الشريان العيني فرع الشريان السباتي الباطن، و يسھم الشريان تحت الحجاجي بجزء من التروية الدموية أيضاً.
فروع الشريان العيني:[33]
- الشريان الشبكي المركزي
- ثمانية شرايين ھدبية خلفية (6 قصيرة و2 طويلة)
- الشرايين الھدبية الأمامية
- فروع عضلية
- الشريان الدمعي
أما العود الوريدي للعين فيصب في الأوردة العينية العلوية والسفلية، استثناء الوريد الشبكي المركزي الذي يصب في الجيب الكھفي (وهنا تبرز خطورة انسداد الوريد الشبكي المركزي لأنه قد يُسبب العمى
صور إضافية
 وصفت هياكل العين
وصفت هياكل العين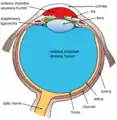 منظر آخر للعين وهياكل العين المسمى
منظر آخر للعين وهياكل العين المسمى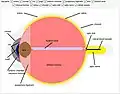 تشريح العين البشرية الذي تم إنشاؤه باستخدام لغة Wolfram
تشريح العين البشرية الذي تم إنشاؤه باستخدام لغة Wolfram
انظر أيضًا
- التكيف (العين) (الرؤية الليلية).
- نظرية الانبعاث (الرؤية).
- لون العين.
- عين الإنسان.
- امراض العين.
- إصابة العين.
- حركة العين.
- جفن.
- الغشاء المضاد.
- طب العيون.
- المدار (علم التشريح).
- عين بسيطة في اللافقاريات.
- بساط شفاف.
- دموع.
- التفاعلات الكيميائية في عملية النظر.
- انشقاق الشبكية.
- التهاب القزحية.
- تطور العين.
- فحص العين.
ملاحظات
- There is no universal consensus on the precise total number of phyla Animalia; the stated figure varies slightly from author to author.
المراجع
اقتباسات
- Land, M.F.; Fernald, R.D. (1992). "The evolution of eyes". Annual Review of Neuroscience. 15: 1–29. doi:10.1146/annurev.ne.15.030192.000245. PMID 1575438. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Frentiu, Francesca D.; Adriana D. Briscoe (2008). "A butterfly eye's view of birds". BioEssays. 30 (11–12): 1151–1162. doi:10.1002/bies.20828. PMID 18937365. الوسيط
|CitationClass=تم تجاهله (مساعدة) - ترجمة و معنى كلمة qiyaamahday في قاموس المعاني. قاموس عربي انجليزي مصطلحات صفحة 1 نسخة محفوظة 26 يونيو 2018 على موقع واي باك مشين.
- Breitmeyer, Bruno (2010). Blindspots: The Many Ways We Cannot See. New York: Oxford University Press. صفحة 4. ISBN 978-0-19-539426-9. مؤرشف من الأصل في 7 مارس 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Nairne, James (2005). Psychology. Belmont: Wadsworth Publishing. ISBN 978-0-495-03150-5. OCLC 61361417. مؤرشف من الأصل في 8 فبراير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bruce, Vicki; Green, Patrick R.; Georgeson, Mark A. (1996). Visual Perception: Physiology, Psychology and Ecology. Psychology Press. صفحة 20. ISBN 978-0-86377-450-8. مؤرشف من الأصل في 8 فبراير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - BioMedia Associates Educational Biology Site: What animal has a more sophisticated eye, Octopus or Insect? نسخة محفوظة 2008-03-05 على موقع واي باك مشين.
- "Who You Callin' "Shrimp"?". National Wildlife Magazine. Nwf.org. 2010-10-01. مؤرشف من الأصل في 22 أبريل 2017. اطلع عليه بتاريخ 03 أبريل 2014. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Meyer-Rochow, V.B. (1974). "Structure and function of the larval eye of the sawfly larva Perga". Journal of Insect Physiology. 20 (8): 1565–1591. doi:10.1016/0022-1910(74)90087-0. PMID 4854430. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cronin, T.W.; Porter, M.L. (2008). "Exceptional Variation on a Common Theme: the Evolution of Crustacean Compound Eyes". Evolution: Education and Outreach. 1 (4): 463–475. doi:10.1007/s12052-008-0085-0. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Kozmik, Z.; Ruzickova, J.; Jonasova, K.; Matsumoto, Y.; Vopalensky, P.; Kozmikova, I.; Strnad, H.; Kawamura, S.; Piatigorsky, J. (2008). "Assembly of the cnidarian camera-type eye from vertebrate-like components". Proceedings of the National Academy of Sciences of the United States of America (Free full text)
|format=بحاجة لـ|url=(مساعدة). 105 (26): 8989–8993. Bibcode:2008PNAS..105.8989K. doi:10.1073/pnas.0800388105. PMID 18577593. الوسيط|CitationClass=تم تجاهله (مساعدة)صيانة CS1: عرض-المؤلفون (link) - Zhukov, ZH; Borisseko, SL; Zieger, MV; Vakoliuk, IA; Meyer-Rochow, VB (2006). "The eye of the freshwater prosobranch gastropod Viviparus viviparus: ultrastructure, electrophysiology and behaviour". Acta Zoologica. 87: 13–24. doi:10.1111/j.1463-6395.2006.00216.x. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Fernald, Russell D. (2006). "Casting a Genetic Light on the Evolution of Eyes" (PDF). Science. 313 (5795): 1914–1918. Bibcode:2006Sci...313.1914F. doi:10.1126/science.1127889. PMID 17008522. مؤرشف من الأصل (PDF) في 28 أكتوبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Circadian Rhythms Fact Sheet". National Institute of General Medical Sciences. مؤرشف من الأصل في 13 مارس 2020. اطلع عليه بتاريخ 03 يونيو 2015. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Kozmik, Zbynek; Ruzickova, Jana; Jonasova, Kristyna; Matsumoto, Yoshifumi; Vopalensky, Pavel; Kozmikova, Iryna; Strnad, Hynek; Kawamura, Shoji; Piatigorsky, Joram (2008). "Assembly of the cnidarian camera-type eye from vertebrate-like components" (PDF). Proceedings of the National Academy of Sciences. 105 (26): 8989–8993. Bibcode:2008PNAS..105.8989K. doi:10.1073/pnas.0800388105. PMC 2449352. PMID 18577593. مؤرشف من الأصل (PDF) في 4 أبريل 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Fernald, Russell D. (2006). "Casting a Genetic Light on the Evolution of Eyes". Science. 313 (5795): 1914–1918. Bibcode:2006Sci...313.1914F. doi:10.1126/science.1127889. PMID 17008522. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Nilsson, Dan-E. (1989). "Vision optics and evolution". BioScience. 39 (5): 298–307. doi:10.2307/1311112. JSTOR 1311112. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Speiser, D.I.; Eernisse, D.J.; Johnsen, S.N. (2011). "A Chiton Uses Aragonite Lenses to Form Images". Current Biology. 21 (8): 665–670. doi:10.1016/j.cub.2011.03.033. PMID 21497091. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wilson, M. (1978). "The functional organisation of locust ocelli". Journal of Comparative Physiology. 124 (4): 297–316. doi:10.1007/BF00661380. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wagner, H.J.; Douglas, R.H.; Frank, T.M.; Roberts, N.W.; Partridge, J.C. (Jan 27, 2009). "A Novel Vertebrate Eye Using Both Refractive and Reflective Optics". Current Biology. 19 (2): 108–114. doi:10.1016/j.cub.2008.11.061. PMID 19110427. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Völkel, R; Eisner, M; Weible, KJ (June 2003). "Miniaturized imaging systems" (PDF). Microelectronic Engineering. 67–68 (1): 461–472. doi:10.1016/S0167-9317(03)00102-3. مؤرشف من الأصل (PDF) في 01 أكتوبر 2008. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Land, Michael (1997). "Visual Acuity in Insects" (PDF). Annual Review of Entomology. 42: 147–177. doi:10.1146/annurev.ento.42.1.147. PMID 15012311. مؤرشف من الأصل (PDF) في 23 نوفمبر 2004. اطلع عليه بتاريخ 27 مايو 2013. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Gaten, Edward (1998). "Optics and phylogeny: is there an insight? The evolution of superposition eyes in the Decapoda (Crustacea)". Contributions to Zoology. 67 (4): 223–236. doi:10.1163/18759866-06704001. مؤرشف من الأصل في 12 يناير 2013. اطلع عليه بتاريخ 11 سبتمبر 2008. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ritchie, Alexander (1985). "Ainiktozoon loganense Scourfield, a protochordate from the Silurian of Scotland". Alcheringa. 9 (2): 137. doi:10.1080/03115518508618961. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Mayer, G. (2006). "Structure and development of onychophoran eyes: What is the ancestral visual organ in arthropods?". Arthropod Structure and Development. 35 (4): 231–245. doi:10.1016/j.asd.2006.06.003. PMID 18089073. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Meyer-Rochow, VB; Gal, J (2004). "Dimensional limits for arthropod eyes with superposition optics". Vision Research. 44 (19): 2213–2223. doi:10.1016/j.visres.2004.04.009. PMID 15208008. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Greiner, Birgit (16 December 2005). Adaptations for nocturnal vision in insect apposition eyes (PDF) (Thesis). Lund University. مؤرشف من الأصل (PDF) في 09 فبراير 2013. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Horváth, Gábor; Clarkson, Euan N.K. (1997). "Survey of modern counterparts of schizochroal trilobite eyes: Structural and functional similarities and differences". Historical Biology. 12 (3–4): 229–263. doi:10.1080/08912969709386565. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Jochen Zeil; Maha M. Al-Mutairi (1996). "Variations in the optical properties of the compound eyes of Uca lactea annulipes" (PDF). The Journal of Experimental Biology. 199 (7): 1569–1577. PMID 9319471. مؤرشف من الأصل (PDF) في 25 فبراير 2009. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Müller, CHG; Rosenberg, J; Richter, S; Meyer-Rochow, VB (2003). "The compound eye of Scutigera coleoptrata (Linnaeus, 1758) (Chilopoda; Notostigmophora): an ultrastructural re-investigation that adds support to the Mandibulata concept". Zoomorphology. 122 (4): 191–209. doi:10.1007/s00435-003-0085-0. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ullrich-Luter, E.M.; Dupont, S.; Arboleda, E.; Hausen, H.; Arnone, M.I. (2011). "Unique system of photoreceptors in sea urchin tube feet". Proceedings of the National Academy of Sciences. 108 (20): 8367–8372. Bibcode:2011PNAS..108.8367U. doi:10.1073/pnas.1018495108. PMID 21536888. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ali & Klyne 1985
- "Anatomy of the Eye". www.hopkinsmedicine.org (باللغة الإنجليزية). مؤرشف من الأصل في 18 سبتمبر 2020. اطلع عليه بتاريخ 03 ديسمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Autrum, H (1979). "Introduction". In H. Autrum (المحرر). Comparative Physiology and Evolution of Vision in Invertebrates- A: Invertebrate Photoreceptors. VII/6A. New York: Springer-Verlag. صفحات 4, 8–9. ISBN 978-3-540-08837-0. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Halder, G.; Callaerts, P.; Gehring, W.J. (1995). "New perspectives on eye evolution". Curr. Opin. Genet. Dev. 5 (5): 602–609. doi:10.1016/0959-437X(95)80029-8. PMID 8664548. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Halder, G.; Callaerts, P.; Gehring, W.J. (1995). "Induction of ectopic eyes by targeted expression of the eyeless gene in Drosophila". Science. 267 (5205): 1788–1792. Bibcode:1995Sci...267.1788H. doi:10.1126/science.7892602. PMID 7892602. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Tomarev, S.I.; Callaerts, P.; Kos, L.; Zinovieva, R.; Halder, G.; Gehring, W.; Piatigorsky, J. (1997). "Squid Pax-6 and eye development". Proc. Natl. Acad. Sci. USA. 94 (6): 2421–2426. Bibcode:1997PNAS...94.2421T. doi:10.1073/pnas.94.6.2421. PMID 9122210. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Conway-Morris, S. (1998). The Crucible of Creation. Oxford: Oxford University Press
- Trevor D. Lamb; Shaun P. Collin; Edward N. Pugh Jr. (2007). "Evolution of the vertebrate eye: opsins, photoreceptors, retina and eye cup". Nature Reviews Neuroscience. 8 (12): 960–976. doi:10.1038/nrn2283. PMID 18026166. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Staaislav I. Tomarev; Rina D. Zinovieva (1988). "Squid major lens polypeptides are homologous to glutathione S-transferases subunits". نيتشر. 336 (6194): 86–88. Bibcode:1988Natur.336...86T. doi:10.1038/336086a0. PMID 3185725. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Eye-Evolution?". Library.thinkquest.org. مؤرشف من الأصل في 15 سبتمبر 2012. اطلع عليه بتاريخ 01 سبتمبر 2012. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Fernald, Russell D. (2001). The Evolution of Eyes: Where Do Lenses Come From? نسخة محفوظة 2006-03-19 على موقع واي باك مشين. Karger Gazette 64: "The Eye in Focus".
- Land, M.F. (1989). "The eyes of hyperiid amphipods: relations of optical structure to depth". Journal of Comparative Physiology A. 164 (6): 751–762. doi:10.1007/BF00616747. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Meyer-Rochow, Victor Benno (1982). "The divided eye of the isopod Glyptonotus antarcticus: effects of unilateral dark adaptation and temperature elevation". Proceedings of the Royal Society of London. B 215 (1201): 433–450. Bibcode:1982RSPSB.215..433M. doi:10.1098/rspb.1982.0052. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Russ, John C. (2006). The Image Processing Handbook. CRC Press. ISBN 978-0-8493-7254-4. OCLC 156223054. مؤرشف من الأصل في 08 فبراير 2021.
The upper limit (finest detail) visible with the human eye is about 50 cycles per degree,... (Fifth Edition, 2007, Page 94)
الوسيط|CitationClass=تم تجاهله (مساعدة) - Klaassen, Curtis D. (2001). Casarett and Doull's Toxicology: The Basic Science of Poisons. McGraw-Hill Professional. ISBN 978-0-07-134721-1. OCLC 47965382. مؤرشف من الأصل في 08 فبراير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The Retina of the Human Eye". hyperphysics.phy-astr.gsu.edu. مؤرشف من الأصل في 14 نوفمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Fischer, Robert E.; Tadic-Galeb, Biljana; Plympton, Rick (2000). Steve Chapman (المحرر). Optical System Design. McGraw-Hill Professional. ISBN 978-0-07-134916-1. OCLC 247851267. مؤرشف من الأصل في 8 فبراير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Barlow, H.B. (1952). "The size of ommatidia in apposition eyes". J Exp Biol. 29 (4): 667–674. مؤرشف من الأصل في 31 أغسطس 2016. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Barlow, Horace Basil; Mollon, J.D. (1982). The Senses. Cambridge: Cambridge University Press. صفحة 98. ISBN 978-0-521-24474-9. مؤرشف من الأصل في 7 مارس 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Fernald, Russell D. (1997). Fernald.pdf "The Evolution of Eyes" تحقق من قيمة
|مسار أرشيف=(مساعدة) (PDF). Brain, Behavior and Evolution. 50 (4): 253–259. doi:10.1159/000113339. PMID 9310200. مؤرشف من الأصل (PDF) في 20 نوفمبر 2012. الوسيط|CitationClass=تم تجاهله (مساعدة) - Goldsmith, T.H. (1990). "Optimization, Constraint, and History in the Evolution of Eyes". The Quarterly Review of Biology. 65 (3): 281–322. doi:10.1086/416840. JSTOR 2832368. PMID 2146698. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Nilsson, E.; Arendt, D. (Dec 2008). "Eye Evolution: the Blurry Beginning". Current Biology. 18 (23): R1096–R1098. doi:10.1016/j.cub.2008.10.025. PMID 19081043. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The Eyes (Human Anatomy): Diagram, Function, Definition, and Eye Problems". WebMD (باللغة الإنجليزية). مؤرشف من الأصل في 30 نوفمبر 2020. اطلع عليه بتاريخ 03 ديسمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة)
قراءة متعمقة
- Yong, Ed (14 January 2016). "Inside the Eye: Nature's Most Exquisite Creation". National Geographic. مؤرشف من الأصل في 01 يونيو 2017. الوسيط
|CitationClass=تم تجاهله (مساعدة)
روابط خارجية
- تطور العين.
- تشريح العين - فلاش متحرك تفاعلي. ( أدوبي فلاش).
- ويبفيجن. تنظيم شبكية العين والجهاز البصري. علاج متعمق لوظيفة الشبكية، مفتوح للجميع ولكنه موجه بشكل أكبر لطلاب الدراسات العليا.
- تظهر أبحاث جامعة كاليفورنيا في بيركلي صورًا لجميع الأساسيات باستثناء الصور الأساسية قبل إرسال المعلومات المرئية إلى الدماغ.
 بوابة طب
بوابة طب بوابة علم الأحياء
بوابة علم الأحياء
