تطور العين
تطور العين هو سلسلة من التطورات التي طرأت لتكوين عين الكائن الحي على مدار ملايين السنين. بعض مكونات العين مثل الأصباغ البصرية تبدو وكأنها من أصل مشترك . وعلى الرغم من أن العين البدائية البسيطة الحساسة للضوء ظهرت فجأة قبل التشعب الحيواني إلا أن العين المعقدة المكونة للصور تطورت حوالي 50 إلى 100 مرة من نفس الجينات و بنفس البروتينات في بناءها، عبر فترة زمنية بدأت في العصر الكامبري قبل نحو 540 مليون سنة.
العين المعقدة تطورت في البداية في بضع ملايين السنين في مرحلة انفجار التطور المعروف بانفجار كامبري. لا يوجد دليل على انه كان يوجد عين قبل العصر الكامبري لكن هناك معدل كبير في التنوع واضح للأحياء وجدت أخفوراتها في طبقات أحجار سجيل صفائحية في سجيل برجس في كولومبيا البريطانية بكندا من الكمبري الوسطى وكذلك وجدت أحافير حيوانات بدائية تبدي أجزاء رخوة متحجرة مثلما في طبقات السجيل في خليج سجيل إيمو باي بجنوب أستراليا . يعود ترسب تلك الطبقات الرسوبية إلى العصر الكامبري الأوسط قبل نحو 505 مليون سنة .
تكيفت العين لتقابل متطلبات المعيشة من طبيعة الأعضاء التي تحملها، وتلعب الجينات في ذلك دورا هاما. يمكن ان تختلف العيون في شكلها وحدتها وكفاءتها، وفي مدي حساسيتها للموجات الضوئية المختلفة، وكذلك بالنسبة لحساسيتها بالنسبة إلى الرؤية بالنهار وفي الظلام أو الليل . ويعتبر قدرة رؤية العين للألوان قد حدث في العصور الجيولوجية الحديثة.
تاريخ الأبحاث
التركيب المعقد للعين استُخدِم كدليل في تدعيم نظرية ان العين صممت بواسطة خالق حيث قيل انه من غير الوارد ان تكون قد تطورت بواسطة الاصطفاء الطبيعي. في سنة 1802 اطلق عليها الفيلسوف ويليام بالي "معجزة تصميم". تشارلز داروين نفسه كتب في كتابه أصل الأنواع ان تطور العين بواسطة الاصطفاء الطبيعي لأول وهلة يبدو "سخيفا" لكنه اخذ في الشرح، أنه بالرغم من صعوبة تخيل الفكرة الا أنها معقولة .
اقترح داروين تدرج من "عصب بصري بالكاد مغلف بالصبغيات دون ان يكون لديه اية آليات أخرى" إلى "درجة أعلى في الكمال" معطيا امثلة لمراحل منقرضة في التطور. اقتراح داروين اكتشف انه صحيح وتحقق الأبحاث الحالية في الآليات الجينية المسئولة عن تطور العين.
في عصرنا الحاضر تطورت أيضا أبحاث التطور وأصبحت تستخدم وسائل في متناولنا الآن وهي التحليل الجيني لدراسة الآلية المسؤولة عن تطور العين والتطور الحيوي بصفة عامة .[2]
سرعة التطور
تعود أقدم الأحافير التي اكتشفت العين حتى الآن إلى الحقبة الكامبيرية (منذ حوالي 540 مليون سنة).[3] شهدت هذه الحقبة تطور سريع يعرف باسم الانفحار الكامبري. واحدة من الفرضيات عن أسباب حدوث هذا التنوع هي نظرية "المبدال الضوئي" لأندرو باركر والتي تقول أن تطور العين بدأ بسباق تسلح والذي أدى إلى حدوث فيض من التطور.[4] وفي المراحل التي سبقت ذلك يمكن ان تكون الأعضاء ذات قد استخدمت مل أجل التنقل السريع عن طريق الرؤية.
ولأن السجل الأحفوري – خاصة في بداية الحقبة الكامبرية – فقير جداً، فإنه من الصعب تقدير سرعة تطور العين. حدوث طفرات صغيرة للنماذج البسيطة وتعرضها للاصطفاء الطبيعي توضح أن عضو الإحساس البصري البدائي المستند إلى أصبغة ضوئية فعالة يمكنه أن يتطور إلى عين معقدة كعين الإنسان في 400.000 سنة تقريباً.[5][هامش 1]
العين في مملكة الحيوان
نحو 95 % من المملكة الحيوانية له أعين . ومن ضمن الحيوانات ذات الأعين يوجد من الفقريات نحو 40.000 نوع، ومن الرخويات و ثنائيات الصدفة و الغمديات نحو 100.000 نوع، ومن مفصليات الأرجل القشريات والعنكبوتيات والحشرات أكثر من مليون نوع. يغلب وجود سلالات الحيوانات ذات أعين في المملكة الحيوانية .[6]
تلك الأغلبية في المملكة الحيوانية ربما ترجع إلى أن تطور العين كان قد بدأ قبل 540 مليون سنة في تريلوبيت ,و ان العين ساعدت على تطور السلالات الحيوانية خلال انفجار الكامبري ؛ في تلك الفترة حدثت تتطورات كثيرة أدى إلى تنوع كبير في المملكة الحيوانية . وطبقا لفرض "أندرو باركر " عالم التاريخ الطبيعي، أن البصر يؤدي إلى توفيق حيوي لواقع العلاقة بين الحيوان المفترس وغنيمته .[7] كما أن البصر يلعب دورا مهما في تعرف الذكور والإناث للجنس اللآخر في مملكة الحيوان.
ويعتبر تطور الحيوانات وزيادة كبرها جسميا خلال تلك المرحلة كان عاملا ضروريا لتطور عينة مركبة . فالشبكية الواسعة المساحة والعدسة الكبيرة والدماغ الكبير الذي به مناطق لمعالجة الإشارات، كل ذلك لا يوجد سوى في حيوانات كبيرة، مثلما تواجدت هذه في العصر الكامبري المبكر في بنيات متعددة مختلفة من الحيوان .[6]
أصل واحد أم عدة أصول؟
لكي نقول أن العين قد تطورت مرة واحدة أو عدة مرات فيتوقف هذا على تعريف العين. معظم الآلية الجينية التي يحدث بها نمو العين مشتركة بين كل الكائنات الحية التي لديها عين، مما يؤدي إلى استنتاج ان سلفهم استخدم نوعا من الية الحساسية للضوء حتى لو لم يكن لديه عضو بصري. لكن حتى الخلايا المستقبلة للضوء يمكن ان تكون قد تطورت أكثر من مرة من مستقبلات كيميائية مشابهة، وأن الخلايا الحساسة للضوء يمكن ان تكون موجودة قبل الانفجار الكامبري بفترة طويلة.[8] الخصائص الأكثر تشابها - مثل استخدام بروتين غلوبولين العدسة في الرأسقدميات وعدسات الفقاريات [9] - تعكس تبدل دور بروتين من دور اولي إلى وظيفة جديدة داخل العين.[10]
مراحل تطور العين
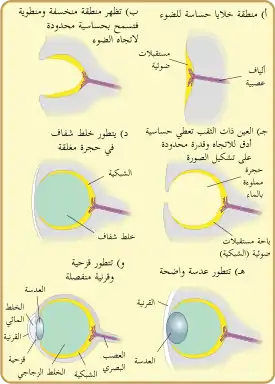
كانت أسلاف العين الأولى المتواجدة لدى الكائنات أحادية الخلية، عبارة عن مستقبلات ضوئية بروتينية حسّاسة للضوء، تسمّى بـ "بقع العين". لا يمكن لبقع العين سوى الإحساس بالسطوع المحيط؛ أي يمكنها التمييز بين الضوء والظلام، وذلك مناسبًا لاستجابة العضويات لطول ساعات النهار (الفترة الضوئية)، وللمزامنة اليومية الخاصة بالنظم اليوماوي. لكنها غير كافية للرؤية، لأنها لا تستطيع التمييز بين الأشكال أو تحديد اتجاه الضوء.
تتواجد بقع العين في جميع المجموعات الحيوانية الرئيسية تقريبًا، وهي شائعة بين الكائنات أحادية الخلية، بما في ذلك الأوغلينا. حيث تقع بقعة الأوغلينا العينية في نهايتها الأمامية، وهي بقعة صغيرة من الصباغ الأحمر الذي يظلل مجموعة من البلورات الحساسة للضوء. ، تسمح البقعة العينية -مع السوط الموجِّه- للكائن الحي بالتحرك استجابةً للضوء، وغالبًا نحو الضوء الذي يساعدها في عملية التمثيل الضوئي، وكذلك في التنبؤ بالليل والنهار -وهو الوظيفة الأساسية للنظم اليوماوي.
تتواجد الأصباغ البصرية في أدمغة الكائنات الأكثر تعقيدًا، ويُعتقَد أنّها تمتلك دورًا في مزامنة التفريخ مع الدورات القمرية. فمن خلال الكشف عن التغييرات الطفيفة في الإضاءة الليلية، يمكن للكائنات الحية مزامنة إطلاق الحيوانات المنوية والبيض لتعزيز احتمال الإخصاب.[11]
تعتمد الرؤية نفسها على الكيمياء الحيوية الأساسية المشتركة بين كل العيون. ومع ذلك، تختلف الطريقة التي تترجم فيها هذه الأدوات الكيميائية الحيوية بيئة العضوية: تحتوي العين على مجموعة واسعة من البُنى والأشكال، التي تطوّرت جميعها بمرحلة متأخرة جدًا مقارنةً بالبروتينات والجزيئات الأساسية.[11]
تمتلك العيون -على المستوى الخلوي- تصميمين رئيسيين؛ أحدهما لدى أوليات الفم (الرخويات، الحلقيات، ومفصليات الأرجل)، والآخر لدى ثانويات الفم (الحبليات، وشوكيات الجلد).
تحتوي الخلية المستقبلة للضوء على بروتينات الأوبسين، وهي الوحدة الوظيفية للعين التي تقوم بإطلاق الإشارة العصبية استجابةً للضوء. تُحمَل الأوبسينات الحسّاسة للضوء على طبقة شعرية، لتعظيم مساحة السطح، وتختلف طبيعة هذه "الشعرات"، حيث يوجد شكلان أساسيان لذلك، وهي: الزغيبات والأهداب. ففي عيون أوليات الفم هي زغيبات: امتدادات أو نتوءات من الغشاء الخلوي، بينما تكون في عيون ثانويات الفم أهدابًا؛ أي بنىً منفصلة. ومع ذلك، قد يستخدم الكائن الحي نوعًا آخرًا من المستقبلات الضوئية خارج العينين، على سبيل المثال: تستخدم الدودة الحلقية (بلاتينيريس ديوميريلياي- Platynereis dumerilii) خلايا زغيبية في العيون، ولكنها تملك أيضًا مستقبلة ضوء هدبية في الدماغ. قد يكون الاشتقاق الفعلي أكثر تعقيدًا، حيث تحتوي بعض الزغيبات على آثارٍ للأهداب. مع ذلك، تدعم الملاحظات الأخرى فرقًا جوهريًا بين أوليات وثانويات الفم. تركّز هذه الاعتبارات على استجابة الخلايا للضوء؛ إذ يستخدم بعضها الصوديوم لإحداث الإشارة الكهربائية التي ستشكل سيالةً عصبيةً، بينما يستخدم البعض الآخر البوتاسيوم. إضافةً إلى ذلك، تقوم أوليات الفم -على العموم- بإحداث الإشارة من خلال السماح للصوديوم بالمرور عبر جدران خلاياها بشكل أكبر، بينما تسمح ثانويات الفم بمرور أقل للصوديوم.[12]
يقترح ذلك امتلاك السلالتين نمطًا بدائيًا من المستقبلات الضوئية، وذلك عندما انفصلتا في العصر ما قبل الكامبري، حيث تطوّرت فيما بعد إلى عيونٍ أكثر تعقيدًا بشكلٍ منفصل.[13]
العيون البدائية
وحدة معالجة الضوء الأساسية للعيون هي الخلية المستقبلة للضوء، وهي خلية متخصّصة تحتوي على نوعين من الجزيئات في الغشاء: أحدهما بروتين حساس للضوء يدعى الأوبسين، والآخر خضاب يميز الألوان يدعى حامل اللون. سُمِّيَت المجموعات المماثلة لهذه الخلايا "بقعًا عينيةً"، ويُعتَقد بأنّها تطورت بشكلٍ مستقلٍ في مكان ما بين 40 و 65 مرة. أتاحت البقع العينية هذه للحيوانات إدراكًا أوليًا جدًا للاتجاه ولشدة الضوء، ولكنها لم تكن كافيةً للتمييز بين الشيء وما يحيطه.
من الواضح أنّ تطوير نظام بصري يمكنه تمييز اتجاه الضوء ضمن بضعة درجات هو أمر أكثر صعوبة بكثير، إذ تمتلك مثل هذا النظام ست شعب فقط من بين ثلاث وثلاثين شعبة. ومع ذلك، تُشَكِّل هذه الشعب 96٪ من الأنواع الحية.[11]
بدأت هذه النظم الضوئية المعقدة بالتزامن مع التقعّر التدريجي للرقع العينية متعددة الخلايا، مما أتاح القدرة على تمييز اتجاهات السطوع، الأمر الذي ازداد دقةً مع زيادة عمق التقعر. لم تكن الرقع العينية المسطحة فعّالة في تحديد اتجاه الضوء، لأنّ الشعاع الضوئي سينشط نفس الخلايا الحساسة للضوء بغض النظر عن اتجاهه، وعلى ذلك، سمح تقعر العيون المجوّفة بتمايز اتجاهي محدود بسبب اختلاف الخلايا التي سينشطها الضوء اعتمادًا على زاويته.[14]
امتلكت الحلزونات القديمة العيون المجوّفة التي ظهرت في العصر الكامبري، ويمكن مشاهدتها اليوم لدى بعض الحلزونات وغيرها من اللافقاريات المعاصرة كالمستورقة. ومع ذلك، كانت هذه العين الأولية مفيدة في تحديد وجود أو غياب الضوء أكثر من تحديدها اتجاهه؛ تغيّر ذلك تدريجيًا مع تعمّق حفرة العين، وتزايد عدد الخلايا المستقبلة للضوء، الأمر الذي زوّد الكائن الحي بمعلوماتٍ بصريةٍ أدق بشكلٍ متزايد.[15]
تسارع تطور العين خلال الانفجار الكامبري بشكل كبير، وشمل ذلك تطورًا جذريًا لكل من معالجة الصور وتحديد اتجاه الضوء.
أصبح دور تقليل عرض فتحة الضوء فيما بعد أكثر فعالية من زيادة عمق التقعر في تحسين الدقة البصرية، إذ وصلت الكائنات الحية عبر ذلك إلى التصوير الحقيقي، الأمر الذي سمح بإدراك دقيق للاتجاهات، إضافةً إلى إدراك قليل للأشكال. تتواجد هذه العيون حاليًا لدى النواتي، لكنها توفر دقة رديئة وتصويرًا خافتًا بسبب افتقارها للقرنية أو العدسة. مع ذلك، لا تزال تُشكّل تحسّنًا كبيرًا مقارنةّ بالعيون البدائية.[16]
مَنَعَ نمو الخلايا الشفافة التلوث والإصابة بالطفيليات، الأمر الذي سمح لمحتويات الحجرة العينية بالتخصص ببطء ضمن خلط شفاف، وبالتالي امتلاك تحسينات كتصفية اللون، معامل الانكسار الأعلى، إعاقة الأشعة فوق البنفسجية، القدرة على العمل داخل وخارج الماء. قد تكون هذه الطبقة مرتبطة بالانسلاخ الذي تمر به بعض الطوائف الحيوانية، كما في حاملات المخالب مثلًا.
تكون القرنية -إضافةً للعدسات والخلطين الشفافين- مسؤولةً عن تجميع الضوء، والمساهمة في تركيزه على الجزء الخلفي لشبكية العين. تحمي القرنية مقلة العين وفي الوقت نفسه تكون مسؤولة عن نحو ثلثي إجمالي قوة الانكسار في العين.[5]
أحد الأسباب الرئيسية المحتملة وراء قدرة العيون على التقاط مجموعة أطوال موجية ضيقة ومحددة من الطيف الكهرومغناطيسي -الطيف المرئي، هو تطوّر الحساسية الضوئية لدى أنواعٍ مائيةٍ في البدء؛ فقط الضوء الأخضر والأزرق المرئيين يمكنهما الانتقال عبر الماء. أثّرت خاصية تصفية الضوء هذه ذاتها على الحساسية الضوئية للنبات.[17]
تطور آلية التركيز
تحرك بعض الأنواع العدسة للخلف والأمام باستخدام عضلة مصرة القزحية، وبعضها يجعل العدسة مسطحة. تنظم آلية أخرى التركيز بشكل كيميائي مستقل عن هذين، عن طريق التحكم في نمو العين والحفاظ على البعد البؤري. إضافةً إلى ذلك، يمكن استخدام شكل البؤبؤ للتنبؤ بالنظام البؤري المُستَخدَم. يمكن أن تشير فرجة البؤبؤ الطولية إلى النظام المشترك متعدد البؤر، بينما يميز البؤبؤ الحلقي عادةً النظام أحادي البؤرة. يتضيق البؤبؤ تحت الضوء الساطع في الشكل الحلقي، مما يزيد البعد البؤري، ويتوسع في الظلام لتقليل عمق التركيز. لاحظ أنّ استخدام آلية التركيز ليست شرطًا. كما يعلم المصورون، تزداد الأخطاء البؤرية مع زيادة الفتحة. وبالتالي، فإنّ عددًا لا يحصى من الكائنات ذات العيون الصغيرة تنشط في ضوء الشمس المباشر، وتعيش بدون آلية تركيز على الإطلاق. لم تظهر وسائل الحاجة للتركيز إلا تدريجيًا، كلما كان النوع أكبر أو عند الانتقال إلى بيئاتٍ أعتم.[18]
مراجع
- Darwin, Charles (1859). أصل الأنواع. London: John Murray.
- Gehring WJ (2005). "New perspectives on eye development and the evolution of eyes and photoreceptors". J. Hered. 96 (3): 171–84. doi:10.1093/jhered/esi027. PMID 15653558. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Parker, A. R. (2009). "On the origin of optics". Optics & Laser Technology. 43 (2): 323–329. Bibcode:2011OptLT..43..323P. doi:10.1016/j.optlastec.2008.12.020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Parker, Andrew (2003). In the Blink of an Eye: How Vision Sparked the Big Bang of Evolution. Cambridge, MA: Perseus Pub. ISBN 0738206075. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Nilsson, D-E; Pelger S (1994). "A pessimistic estimate of the time required for an eye to evolve". Proc R Soc Lond B. 256 (1345): 53–58. doi:10.1098/rspb.1994.0048. PMID 8008757. الوسيط
|CitationClass=تم تجاهله (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Lane, Nick. Leben. Verblüffende Erfindungen der Evolution. Primus Verlag 2013
- Parker, Andrew . In the Blink of an Eye. Free Press 2003
- Nilsson, D. E. (1996). "Eye ancestry: old genes for new eyes". Current biology : CB. 6 (1): 39–42. doi:10.1016/S0960-9822(02)00417-7. PMID 8805210. الوسيط
|CitationClass=تم تجاهله (مساعدة) - doi:10.1016/S0167-4838(99)00066-7
- PMID 20394064 (ببمد 20394064)
Citation will be completed automatically in a few minutes. Jump the queue or expand by hand -
M F Land; R D Fernald (1992). "The Evolution of Eyes". Annual Review of Neuroscience. 15: 1–29. doi:10.1146/annurev.ne.15.030192.000245. PMID 1575438. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Autrum, H (1979). "Introduction". In H. Autrum (المحرر). Comparative Physiology and Evolution of Vision in Invertebrates- A: Invertebrate Photoreceptors. VII/6A. New York: Springer-Verlag. صفحات 6–9. ISBN 978-3-540-08837-0. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Arendt, D.; Tessmar-Raible, K.; Snyman, H.; Dorresteijn, A.W.; Wittbrodt, J. (29 October 2004). "Ciliary Photoreceptors with a Vertebrate-Type Opsin in an Invertebrate Brain". Science. 306 (5697): 869–871. Bibcode:2004Sci...306..869A. doi:10.1126/science.1099955. PMID 15514158. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Fernald, Russell D. (2001). The Evolution of Eyes: How Do Eyes Capture Photons? نسخة محفوظة 19 March 2006 على موقع واي باك مشين. Karger Gazette 64: "The Eye in Focus".
- Eye-Evolution? نسخة محفوظة 15 September 2012 على موقع واي باك مشين.
- Gehring, W. J. (13 January 2005). "New Perspectives on Eye Development and the Evolution of Eyes and Photoreceptors". Journal of Heredity. 96 (3): 171–184. doi:10.1093/jhered/esi027. PMID 15653558. مؤرشف من الأصل في 10 مايو 2010. اطلع عليه بتاريخ 26 أبريل 2008. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ali, M.A. and M. A. Klyne. 1985. Vision in vertebrates. New York: Plenum Press
- Schoenemann, B.; Liu, J. N.; Shu, D. G.; Han, J.; Zhang, Z. F. (2008). "A miniscule optimized visual system in the Lower Cambrian". Lethaia. 42 (3): 265–273. doi:10.1111/j.1502-3931.2008.00138.x. الوسيط
|CitationClass=تم تجاهله (مساعدة)
هوامش
- شكك ديفيد برلينسكي، وهو من أنصار التصميم العاقل، في طريقة الحسابات، وقد دحض مؤلف المقالة الأصلية انتقاداته. انظر:نسخة محفوظة 25 نوفمبر 2017 على موقع واي باك مشين.
- Berlinski, David (2001). "Commentary magazine". مؤرشف من الأصل في 20 يوليو 2017. الوسيط
|CitationClass=تم تجاهله (مساعدة); Cite journal requires|journal=(مساعدة) - Nilsson, Dan-E. "Beware of Pseudo-science: a response to David Berlinski's attack on my calculation of how long it takes for an eye to evolve". مؤرشف من الأصل في 21 مايو 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة); Cite journal requires|journal=(مساعدة) - "Evolution of the Eye" on PBS
- Berlinski, David (2001). "Commentary magazine". مؤرشف من الأصل في 20 يوليو 2017. الوسيط
