جهاز الرؤية
جهاز الرؤية هو الجزء من الجهاز العصبي المركزي الذي يمكن الكائنات من معالجة التفاصيل المرئية، وكذلك تمكين العديد من الدالات الاستجابة المرئية من تشكيل الصورة. يقوم هذا الجهاز بتفسير المعلومات من الضوء المرئي لبناء تمثيل للعالم المحيط. يقوم جهاز الرؤية بإنجاز عدد من المهام المعقدة، بما في ذلك استقبال الضوء وتشكيل تمثيلات الألوان، وبناء تصور منظوري من زوج الإسقاطات ثنائية الأبعاد، وتحديد وتصنيف الكائنات المرئية، وتقييم المسافات عن وبين الكائنات، وتوجيه حركات الجسم بالنسبة للكائنات المرئية. تعرف العملية النفسية لإظهار المعلومات المرئية باسم التصور المرئي، وقلته يؤدي إلى ما يعرف باسم العمى.
مقدمة
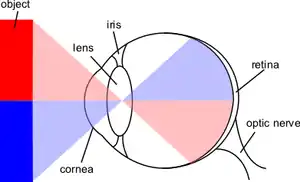
تتحدث هذه المقالة في معظمها عن جهاز الرؤية في الثدييات. يتكون جهاز الرؤية عادة من:
- العين، ولا سيما الشبكية
- العصب البصري
- التصالب البصري
- المسالك البصرية
- الجسم الوحشي الركبي
- الإشعاع البصري
- القشرة البصرية
- قشرة الرابطة البصرية.
تكون الأنواع المختلفة قادرة على رؤية أجزاء مختلفة من الطيف الضوئي، على سبيل المثال، يمكن أن ترى نحلة إلى الطيف في الأشعة فوق البنفسجية، [1] بينما يمكن لأفعى الحفرة إيجاد الفريسة المستهدفة مع باستخدام جهاز الحفرة الذي تستخدمه لتحسس الأشعة تحت الحمراء.[2]
نبذة عن الجهاز
ميكانيكيًا
تعكس العدسة والقرنية معًا الضوء كصورة صغيرة تلمع على الشبكية. ثم تنقل الشبكية هذه الصورة على شكل نبضات كهربائية بواسطة العصي والمخاريط، ثم يحمل العصب البصري هذه النبضات عن طريق السبيل البصري. تتشابك الألياف البصرية عند بلوغ العصب البصري (اليمين يصبح يسار). ثم تتفرع الألياف وتنتهي في ثلاثة أماكن.[3][4][5][5][6][7][8]
عصبيًا
تنتهي معظم ألياف العصب البصري في النَّواة الرُّكْبيَّة الوَحشِيَّة. قبل أن توجّه النَّواةُ الرُّكْبِيَّةُ الوَحْشِيَّة النبضات إلى القشرة البصرية الأولى في الباحة البصرية (الأولية) فإنها تقيس مجال العناصر وتميز كل عنصر بعلامة سرعة. تتوقع هذه العلامات حركة العناصر. وترسل النواة الركبية الوحشية بعض الألياف العصبية إلى القشرتان البصريتان الثانية والثالثة.[9][10][11][12][13]
تتحرى القشرة البصرية الأولى الحواف لفهم التنظيم الحَيِّزِيّ (في البداية خلال 40 ميلي ثانية تُركِّز حتى على الأحياز الصغيرة وتغيرات الألوان، عند تلقي المعلومات المفسرة من قبل النواة الركبية الوحشية والقشرة البصرية الثالثة تبدأ بالتركيز أيضًا على التنظيم الشامل).
تقوم القشرة البصرية الثانية بكل من نقل (مباشرةً وعن طريق وسادة المهاد) النبضات إلى القشرة البصرية الأولى وتلقيهم. وسادة المهاد مسؤولة عن الرَمْش والانتباه البصري. تقوم القشرة البصرية الثانية بنفس وظيفة القشرة البصرية الأولى تقريباَ، وتتعامل أيضًا مع الأشكال الوهمية محددةً العمق من خلال مقارنة النبضات اليمنى واليسرى (صورة ثنائية الأبعاد) ومع التمييز الأمامي. تتصل القشرة البصرية الثانية بكل من القشرتين البصريتين الخامسة والأولى.
تساعد القشرة البصرية الثالثة بمعالجة حركة العناصر العامة (الاتجاه والسرعة). تتصل القشرة البصرية الثالثة بالقشرة البصرية الأولى (بشكل ضعيف) وبالثانية وبالباحة الصُّدْغِيُّة السُّفْلِيّة.[14][15]
تميز القشرة البصرية الرابعة الأشكال البسيطة وتتلقى الإشارات من القشرة البصرية الأولى (بشكل قوي) والثانية والثالثة ومن النواة الركبية الوحشية ومن وسادة المهاد. تشمل الإشارات الصادرة عن القشرة البصرية الخامسة القشرة البصرية الرابعة والمنطقة المحيطة بها والقشور الحركية لحركة العين (مجال العين الأمامي والمنطقة داخل الجدارية الوحشية).[16]
وظيفة القشرة البصرية الخامسة مشابهة للقشور البصرية الأخرى ولكنها تدمج حركة العناصر القريبة ضمن الحركة العامة على درجة من التعقيد. تعمل القشرة البصرية السادسة مع الخامسة على تحليل الحركة. تحلل القشرة البصرية الخامسة الحركة الذاتية، في حين تحلل القشرة البصرية السادسة حركة العناصر بالنسبة للخلفية. ترسل القشرة البصرية السادسة إشاراتها الأولية إلى القشرة البصرية الأولى بالإضافة إلى الإشارات الواردة من القشرة البصرية الخامسة. تحتوي القشرة البصرية السادسة على الخريطة الطبوغرافية للرؤية. تصدر الإشارات عن القشرة البصرية السادسة إلى المنطقة المحيطة بها مباشرةً وتسمى القشرة البصرية السادسة أ. يوجد صلة مباشرة بين القشرة البصرية السادسة أ والباحة المسؤولة عن تحريك اليد بما فيها الباحة أَمامَ الحَرَكِيَّة.[17][18]
يميز التَّلْفيفُ الصُّدْغِيُّ السُّفْلِيّ الأشكال المعقدة والعناصر والوجوه أو، بالترابط مع الحصين، يخلق ذكريات جديدة.[19] تتألف الباحَةُ أَمامَ السَّقْفِيَّة من سبع نوى فريدة. تُثبط النوى أمام السقفية الأمامية والخلفية والأنسية الألم (بشكل غير مباشر) وتساعد في الحَرَكات العَينيَّة السريعة والتكيُّف الانعكاسيّ على التوالي. تعدل نَواةُ إيدنغَر-ويستفال (نَواةُ مُحَرِّكِ العَينِ الإِضافِيَّة) توسع الحدقة وتساعد (نظرًا لأنها تمد بالألياف نظيرة الودية) في تقارب العينين وملائمة العدسة.[20][21]
تشارك نوى السبيل البصري في حركات الملاحقة السلسة التي تقوم بها العين والتكيف الانعكاسي وكذلك الحَرَكاتٌ العَينِيَّةٌ السَريعَة.
النَّواة فوق التصالب هي منطقة من الحصين توقف إنتاج الميلاتونين (بشكل غير مباشر) عند الضوء الأول.[22]
البنية
- المقلة وخصوصًا الشبكية
- العصب البصري
- التصالب البصري
- السبيل البصري
- النَّواةُ الرُّكْبِيَّةُ الوَحْشِيَّة
- التشعع البصري
- القشرة البصرية
- القشرة البصرية المشاركة
كل من هذه مقسمة إلى سبل أمامية وخلفية. يشير السبيل البصري الأمامي إلى البنى المشاركة في الرؤية قبل النواة الركبية الوحشية في حين تشير السبل الخلفية إلى البنى المشاركة في الرؤية بعد هذه النقطة.
المقلة
ينعكس الضوء الذي يدخل إلى العين عند مروره عبر القرنية. ثم يعبر الحدقة (التي تتحكم بها القزحية) ثم ينكسر أكثر بواسطة العدسة. تعمل القرنية والعدسة كعدسة واحدة بهدف عرض صورة مقلوبة على الشبكية.
الشبكية
تتكون البشكية من عدد كبير من الخلايا المستقبلة للضوء التي تحتوي على جزيئات بروتينية معينة تسمى الأوبسينات. هناك نوعان من الأوسينات التي تساهم في الرؤية الواعية لدى الإنسان: أوبسينات العصي وأوبسينات المخاريط (هناك نواع ثالث وهو الميلانوبسين في بعض الخلايا العقدية في الشبكية) وهي جزء من آلية ساعة الجسم وغالبًا ليس لها علاقة بالرؤية الواعية نظرًا لأنها غير متصلة بالنواة الركبية الوحشية بل بالنواة الزيتونية أمام السقفية. يمتص الأوبسين الفوتونات (الجزيئات الضوئية) وتنقل الإشارة على الخلية عبر سبيل تحويل الإشارة ما يؤدي إلى فرط استقطاب في المستقبل الضوئي.[23]
تختلف العصي والمخاريط وظيفيًا. تتواجد العصي في المقام الأول في محيط شبكية العين وتُستخدم للرؤية ضمن الإضاءة المنخفضة. تتواجد المخاريط في المقام الأول في مركز (اللطخة الصفراء) الشبكية. هناك ثلاثة أنواع للمخاريط وتختلف في طول الموجة الضوئية التي تمتصها والتي تسمى عادةً القصيرة أو الزرقاء، المتوسطة أو الخضراء والطويلة أو الحمراء. تستخدم المخاريط في المقام الأول بتمييز الألوان والمعالم الأخرى ضمن المحيط البصري وضمن مستويات طبيعية من الإضاءة.[24]
تتشابك المستقبلات الضوئية في الشبكية بشكل مباشر مع الخلايا ثنائية القطب والتي بدورها تتشابك مع الخلايا العقدية في الطبقة الخارجية والتي ستولد بعد ذلك إمكانيات التنبيه في الدماغ. ينشأ قدر كبير من المعالجة البصرية من أنماط التواصل بين الخلايا العصبية في شبكية العين. هناك نحو 130 مليون مستقبل ضوئي يمتص الضوء ولكن ما يقارب 1.2 مليون محور عصبي من الخلايا العقدية تنقل المعلومات من شبكية العين إلى الدماغ. تشمل المعالجة في الشبكية تشكيل حقول استقبال حول مركزية من الخلايا ثنائية القطب والعقدية الشبكية وكذلك التقارب والتباعد من المستقبل الضوئي إلى الخلايا ثنائية القطب. بالإضافة إلى ذلك فإن الخلايا العصبية الأخرى في الشبكية، وخصوصًا الخلايا الأفقية والخلايا عديمة المحور، تنقل المعلومات بشكل وحشي (من الخلايا العصبية في طبقة إلى الخلايا العصبية الأخرى في نفس الطبقة) ما يؤدي إلى حقول استقبال أكثر تعقيدًا تكون إما غير قادرة على تمييز الألوان وقادرة على تمييز الحركة أو العكس.[25]
آلية توليد الإشارات البصرية: تتكيف الشبكية مع تغيرات الضوء باستخدام العصي. يأخذ حامل اللون الشبكي في الظلام شكل منحنى يسمى المقرون الشبكي. عندما يتفاعل الضوء مع الشبكية يتغير التشكل إلى شكل مستقيم يسمى العابر للشبكية وينفصل عن الأوبسين. هذا يسمى التبييض لأن الرودوبسين المنقى يتغير من البنفسجي إلى عديم اللون. في الأساس في الظلام لا يمتص الرودوبسين أي ضوء ويطلق الغلوتامات الذي يثبط الخلية ثنائية القطب. هذا يثبط تحرير النواقل العصبية من الخلايا ثنائية القطب إلى الخلايا العقدية. في حالة وجود الضوء فإن إفراز الغلوتامات ينخفض وبالتالي لا يوجد تثبيط على إطلاق الخلايا ثنائية القطب للنواقل العصبية إلى الخلايا العقدية وبالتالي يُكتشف وجود الصورة.[26][27]
انظر أيضاً
مراجع
- Bellingham et al. 1997، صفحات 775–781
- Safer & Grace 2004، صفحات 55–61.
- "How the Human Eye Sees." WebMD. Ed. Alan Kozarsky. WebMD, 3 October 2015. Web. 27 March 2016.
- Than, Ker. "How the Human Eye Works." LiveScience. TechMedia Network, 10 February 2010. Web. 27 March 2016.
- "How the Human Eye Works | Cornea Layers/Role | Light Rays." NKCF. The Gavin Herbert Eye Institute. Web. 27 March 2016.
- Tillotson, Joanne. McCann, Stephanie. Kaplan’s Medical Flashcards. April 2, 2013.
- "Optic Chiasma." Optic Chiasm Function, Anatomy & Definition. Healthline Medical Team, 9 March 2015. Web. 27 March 2016.
- Jefferey, G., and M. M. Neveu. "Chiasm Formation in Man Is Fundamentally Different from That in the Mouse." Nature.com. Nature Publishing Group, 21 March 2007. Web. 27 March 2016.
- Card, J. Patrick, and Robert Y. Moore. "Organization of Lateral Geniculate-hypothalamic Connections in the Rat." Wiley Online Library. 1 June. 1989. Web. 27 March 2016.
- Murphy, Penelope C., Simon G. Duckett, and Adam M. Sillito. "Feedback Connections to the Lateral Geniculate Nucleus and Cortical Response Properties." Feedback Connections to the Lateral Geniculate Nucleus and Cortical Response Properties. 19 November 1999. Web. 27 March 2016.
- Schiller, P. H., and J. G. Malpeli. "Functional Specificity of Lateral Geniculate Nucleus Laminae of the Rhesus Monkey." APS Journals. 1 May 1978. Web. 27 March 2016.
- Singer, W., and F. Schmielau. "The Role of Visual Cortex for Binocular Interactions in the Cat Lateral Geniculate Nucleus." The Role of Visual Cortex for Binocular Interactions in the Cat Lateral Geniculate Nucleus. 21 January 1977. Web. 27 March 2016.
- Reed, R. Clay, and Jose-Manuel Alonso. "Specificity of Monosynaptic Connections from Thalamus to Visual Cortex." Letters to Nature. Nature Publishing Group, 3 October 1995. Web. 27 March 2016.
- Heim, Stefan, Simon B. Eickhoff, et al. "Effective Connectivity of the Left BA 44, BA 45, and Inferior Temporal Gyrus during Lexical and Phonological Decisions Identified with DCM." Wiley Online Library. 19 December 2007. Web. 27 March 2016.
- Catani, Marco, and Derek K. Jones. "Brain." Occipito‐temporal Connections in the Human Brain. 23 June 2003. Web. 27 March 2016.
- Benevento, Louis A., and Gregg P. Strandage. "The Organization of Projections of the Retinorecipient and Nonretinorecipient Nuclei of the Pretectal Complex and Layers of the Superior Colliculus to the Lateral Pulvinar and Medial Pulvinar in the Macaque Monkey." Science Direct. 1 July 1983. Web. 27 March 2016.
- Hirsch, JA, and CD Gilbert. "The Journal of NeuroscienceSociety for Neuroscience." Synaptic Physiology of Horizontal Connections in the Cat's Visual Cortex. 1 June 1991. Web. 27 March 2016.
- Schall, JD, A. Morel, DJ King, and J. Bullier. "The Journal of NeuroscienceSociety for Neuroscience." Topography of Visual Cortex Connections with Frontal Eye Field in Macaque: Convergence and Segregation of Processing Streams. 1 June 1995. Web. 27 March 2016.
- Moser, May-Britt, and Edvard I. Moser. "Functional Differentiation in the Hippocampus." Wiley Online Library. 1998. Web. 27 March 2016.
- Kanaseki, T., and J. M. Sprague. "Anatomical Organization of Pretectal Nuclei and Tectal Laminae in the Cat." Anatomical Organization of Pretectal Nuclei and Tectal Laminae in the Cat. 1 December 1974. Web. 27 March 2016.
- Reiner, Anton, and Harvey J. Karten. "Parasympathetic Ocular Control — Functional Subdivisions and Circuitry of the Avian Nucleus of Edinger-Westphal."Science Direct. 1983. Web. 27 March 2016.
- Welsh, David K., and Diomedes E. Logothetis. "Individual Neurons Dissociated from Rat Suprachiasmatic Nucleus Express Independently Phased Circadian Firing Rhythms." Science Direct. Harvard University, April 1995. Web. 27 March 2016.
- Güler, A.D.; et al. (May 2008). "Melanopsin cells are the principal conduits for rod/cone input to non-image forming vision" (Abstract). Nature. 453 (7191): 102–5. Bibcode:2008Natur.453..102G. doi:10.1038/nature06829. PMC 2871301. PMID 18432195. الوسيط
|CitationClass=تم تجاهله (مساعدة)CS1 maint: ref=harv (link) - Nave, R. "Light and Vision". HyperPhysics. مؤرشف من الأصل في 14 أكتوبر 2019. اطلع عليه بتاريخ 13 نوفمبر 2014. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Tovée 2008
- Saladin, Kenneth D. Anatomy & Physiology: The Unity of Form and Function. 5th ed. New York: McGraw-Hill, 2010.
- "Archived copy". مؤرشف من الأصل في 23 يناير 2011. اطلع عليه بتاريخ 08 ديسمبر 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة)صيانة CS1: الأرشيف كعنوان (link)
 بوابة تشريح
بوابة تشريح بوابة طب
بوابة طب بوابة علم وظائف الأعضاء
بوابة علم وظائف الأعضاء بوابة علوم عصبية
بوابة علوم عصبية
