عائلة راس
عائلة راس[1] عائلة بروتينات معربة في جميع خلايا أعضاء الحيوانات. جميع بروتينات هذه العائلة تنتمي إلى فئة من البروتين تسمى GTPase الصغيرة (بالإنجليزية:small GTPase)، وتشترك في نقل الإشارات داخل الخلية (التنبيغ الخلوي). راس هي العضو النموذجي في فصيلة راس (بالإنجليزية:Ras superfamily)، حيث أن أعضاء هذه الفصيلة تتشابه في الهيكل ثلاثي الأبعاد وتقوم بتنظيم سلوكات ووظائف متنوعة داخل الخلية.
 | |
| معرف | |
| رمز | Ras |
| قاعدة بيانات عوائل البروتينات | PF00071 |
| إنتربرو | IPR020849 |
| بروسايت | PDOC00017 |
| قاعدة بيانات التصنيف الهيكلي للبروتينات | 5p21 |
بعد أن يتم تشغيل البروتين راس بواسطة الإشارات الواردة، يقوم راس بتشغيل بروتين آخر، والذي بدوره يقوم بتشغيل جينات، حيث أن هذه الجينات لها دور في عمليات: تكاثر الخلايا، وتمايزها، بالإضافة إلى بقائها على قيد الحياة.
تؤدي الطفرات التي تصيب الجينات المترجمة لبروتينات راس إلى إنتاج بروتين راس دائم النشاط والفاعلية. نتيجة لذلك، تتكون إشارات غير مقصودة وشديدة النشاط داخل الخلية، حتى في حالة غياب إشارات واردة من خارج الخلية تحفز تكونها داخل الخلية. وبما أن هذه الإشارات تؤدي إلى نمو الخلية وانقسامها، فالنشاط الشديد لإشارات راس سوف تؤدي إلى نمو غير طبيعي للخلية الذي ينتج عنه تكون الورم السرطاني.[2]
تحتوي خلايا الإنسان على ثلاث جينات مترجِمة لبروتينات راس تسمى "جينات راس الثلاث" وهي:KRAS ،HRAS، وNRAS. تعد هذه الجينات من أكثر الجينات المسرطنة شيوعاً، من خلال الطفرات التي تسبب نشاط دائم لبروتينات راس. تتواجد هذه الطفرات من 20% إلى 25% من مجموع الأورام التي تصيب الإنسان، وفي أنواع معينة من السرطان تتواجد بنسبة 90% (مثل سرطان البنكرياس).[3] ولهذا السبب، يتم دراسة مثبطات راس لاستخدامها كعلاج للسرطان وأمراض أخرى تتصف بإنتاج مفرط لبروتينات راس.
تاريخ
تم اكتشاف أول جينَين مترجمَين لبروتنيات راس (HRAS وKRAS)[4] من خلال الدراسات التي أُجريت على نوعين من الفيروسات المسببة للسرطان، هما: فيروس هارفي وفيروس كريستن، من قبل إدوارد م.سكولنك وزملاؤه في منظمات الصحة الوطنية (National Institutes of Health).[5] لقد اكتُشفت هذه الفيروسات داخل الفئران في السيتينات من القرن الماضي من قبل جينيفر هارفي[6] وويرنر كريستن،[7] على التوالي، وعليه سمي بورم الفئران الخبيث (ساركوما الفئران) .[4]
عام 1982 جينات راس المفعلة والمحولة اكتشفت في جسم الإنسان داخل الخلايا السرطانية من قبل جيفري م. كوبر في هارفرد،[8] ماريانو بارباسيد وستيوارت أ. آرونسون في منظمات الصحة الوطنية،[9] روبرت واينبرغ في معهد ماساتشوستس للتكنولوجيا،[10] ومايكل ويغلر في مختبر كولد سبرينغ هاربور.[11] بعد ذلك تم التعرف على الجين الثالث على يد مجموعة من الباحثين : روبن فايس في معهد أبحاث السرطان،[12][13] ومايكل ويغلر في مختبر كولد سبرينغ هاربور،[14] سمي هذا الجين بNRAS لدوره المهم في تحديد طبيعة الورم الخبيث الذي يصيب الخلايا العصبية البشرية (Neuroblastoma).
جينات راس الثلاثة تشفر بروتينات في غاية التشابه، تتكون من سلاسل من 188 إلى 189 حمض أميني. رموز الجينات الخاصة بهذه البروتينات هي: HRAS، KRAS، و NRAS.KRAS ينتج النظائر K-Ras4A و K-Ras4B من خلال الروابط التجديلية.
بنية بروتينات راس
يحتوي البروتين على ستة خيوط بيتا (بالانجليزية: beta strands) وخمسة لوالب الفا (بالانجليزية:alpha helices) [15] كما يتركب من مجموعتين رئيسيتين، المجموعة (ج) تتكون من 166 حمض اميني، ما يقارب 20 كيلودالتون، الذي يربط النوكليوتيدات الغوانوزينية، ومنطقة استهداف الغشاءالطرفي (سي) (CAAX-COOH )، المعروف أيضًا باسم ( CAAX box)، وهو معدل للدهون بواسطة فرانسيل ترانسفيراز ( بالانجليزية: farnesyl transferase) و RCE1 و ICMT.
تحتوي المجموعة (ج) على خمسة مجموعات جزئية من (ج) التي تربط الناتج المحلي الإجمالي GTP مباشرة.
العنصر (ج1) أو P-loop، إلى فوسفات بيتا من الناتج المحلي الإجمالي و GTP.العنصر(ج2) والمعروف أيضاً بالمفتاح 1، يحتوي على ثريونين 35، الذي يربط الفوسفات الطرفي (γ-phosphate) لـ GTP وأيون المغنيسيوم ثنائي التكافؤ المتواجد في الموقع النشط.العنصر (ج3)، الذي يُعرف أيضًا بـالمفتاح 2 ،يحتوي ( DXXGQ ) , (D) هو الاسبرتيت 57 ، وهو محدد لربط الغوانين مقابل الأدينين، و (Q) هو جلوتامين 61 ، وهو المتبقي الحيوي الذي ينشط جزيء ماء محفز للتحلل بالماء من (GTP) إلى إجمالي الناتج المحلي
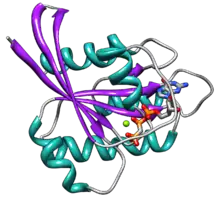
العنصر(ج4) يحتوي على نموذج (LVGNKxDL) ، ويوفر تفاعلًا محددًا مع الجوانين.
يحتوي العنصر (ج5) على تسلسل إجماع (SAK. A) وهو الانيين 146،الذي يوفر خصوصية لغوانين بدلا من الأدينين.
يمثل كل من المفتاح 1 والمفتاح 2 ، (ج2)و(ج3) ، الأجزاء الرئيسية من البروتين التي تتحرك عند التنشيط بواسطة GTP.
هذا التغير التطابقي من خلال (ج2) و (ج3) هو ما يتوسط الوظيفة الأساسية كبروتين تبديل جزيئي.
حالة بروتين راس القائمة على بروتوكول GTP هي حالة "التشغيل"، والحالة المرتبطة بالناتج المحلي الإجمالي (GDP) هي حالة "التعطيل".
بروتين راس أيضا يحتوي [أيون المغنيسيوم] الذي يساعد على تنسيق ربط النوكليوتيدات.
الوظيفة
تعمل بروتينات راس كمفاتيح جزيئية ثنائية تتحكم في شبكات الإشارات داخل الخلايا. تتحكم بمسارات توصيل الاشارة للتنظيم في مثل هذه العمليات كسلامة الهيكل الخلوي للاكتين، وتكاثر الخلايا، وتمايز الخلايا، والتصاق الخلايا، واستماتة الخلية، وترحيل الخلايا (بالإنجليزية: cell migration). غالبًا ما يتم تحرير البروتينات ذات الصلة بالبروتين راس وبروتين راس في السرطانات، مما يؤدي إلى زيادة الغزو والانبثاث، وتراجع الموت المنظم للخلية.
ينشط البروتين راس العديد من المسارات، التي تمت دراستها بشكل جيد. ينقل هذا البروتين الإشارات باتجاه محدد وينتج عنه نسخ الجينات المعنية بنمو الخلايا وتقسيمها . وينقل البروتين راس اشارات تنشيط مسار آخر هو مسار (PI3K/AKT/mTOR pathway)، الذي يحفز تخليق البروتين والنمو الخلوي، ويحول دون موت الخلايا المنظم.
التنشيط والغاء التنشيط
البروتين راس هو جي بروتين، أو بروتين ربط غوانوزين - النوكليوتيد. على وجه التحديد، وهو عبارة عن وحدة (بروتين الGTPase) وهي وحدة صغيرة أحادية، وهو مرتبط في بنية الوحدة الفرعية (جα).
تعمل البروتينات (ج) كمفاتيح تشوير ثنائية مع حالات "التشغيل" و"ايقاف التشغيل". في حالة "إيقاف التشغيل"، يكون مرتبطًا بنواة ثنائي أكسيد الجوانوزين (بالإنجليزية: guanosine diphosphate (GDP).
بينما في حالة "التشغيل"، يكون مرتبطا بغوانوسين ثلاثي الفوسفات (بالانجليزية : (guanosine triphosphate (GTP)، التي لديها مجموعة فوسفات إضافية مقارنة بالناتج المحلي الإجمالي (GDP ).
يحمل هذا الفوسفات الإضافي منطقتين للتبديل في تكوين "الربيع المحمل" (على وجه التحديد، Thr-35 و Gly-60). مناطق تبديل الاسترخاء الذي يسبب تغيير في الحالة إلى غير نشط. وبالتالي، يتم التحكم في تنشيط وتعطيل راس وغيرها من البروتينات (ج) الصغيرة عن طريق التدوير بين الحالتين في (GTP) والحالة الناتجة عن الناتج المحلي الإجمالي(GDP).
يتم تسهيل عملية تبادل النوكليوتيدات المربوطة من قبل عوامل تبادل النوكليوتيدات في الغوانين (GEF), و GTPase لتنشيط البروتينات (GAPs). حسب تصنيفها، فإن بروتين راس لها نشاط بروتين الGTPase جوهري، ما يعني أن البروتين بمفرده يقوم بتحلل مائي لجزيء GTP إلى إجمالي الناتج المحلي(GDP). ومع ذلك، فإن هذه العملية بطيئة للغاية بالنسبة للوظيفة الفعالة، وبالتالي فإن GAP في راس، RasGAP ، قد ترتبط وتثبّت الماكينة التحفيزية في بروتين راس، توفير بقايا تحفزية إضافية ("إصبع أرجينين") بحيث يتم وضع جزيء الماء على النحو المحب للنواة على فوسفات جاما من GTP. يتم تحرير الفوسفات غير العضوي ويكون جزيء الراس مرتبطًا بالناتج المحلي الإجمالي. ونظرًا لأن الشكل المحدد للناتج المحلي الإجمالي هو "إيقاف" أو "غير نشط" للإشارة، فإن GTPase Activating Protein يعطل رأسه عن طريق تنشيط نشاط GTPase.
وبالتالي، تعمل GAP على تسريع تعطيل نشاط بروتين راس.
تقوم GEF بتحفيز رد فعل "الدفع والجذب" الذي يطلق الناتج المحلي الإجمالي من بروتين راس. أنها تضاف قريبة من موقع ربط حلقة بي وايون المغنيسيوم وتمنع تفاعل هذه مع أيون فوسفات جاما. البقايا الحمضية (السلبية) في التبديل II "سحب" ليسين في P-lo بعيدا عن الناتج المحلي الإجمالي الذي "يدفع" التبديل بعيدا عن الجوانين. تم كسر جهات الاتصال التي تحتفظ بالناتج المحلي الإجمالي في المكان ويتم إطلاقها في السيتوبلازم.
لأن GTP داخل الخلايا وفيرة نسبة إلى الناتج المحلي الإجمالي (حوالياكثر من 10 أضعاف) [15] GTP في الغالب يقوم بإعادة إدخال جيب الربط النوكليوتيد من البروتبن راس وإعادة تحميل الربيع.
وبالتالي فإن GEF تسهل تفعيل رأس.[15] وتشمل GEFs المعروفة جيدا (Son of Sevenless) و cdc25 التي تتضمن نطاق RasGEF.
يحدد التوازن بين نشاط GEF و GAP وضع النوكليوتيد الغوانين في البروتين راس، وبالتالي تنظيم نشاط البروتين راس.
في التصميم المكوّن من GTP ، فإن راس لديها تقارب كبير للعديد من المستجيبات التي تسمح لها بتنفيذ وظائفها. وتشمل هذه PI3K. GTPases صغيرة أخرى لربط محولات مثل ارفبتين أو أنظمة رسول الثاني مثل انزيم محلقة الأدينيلات.
تم العثور على المجموعة الرابطة راس في العديد من المستجيبات ويربط دائمًا بإحدى مناطق التحويل، نظرًا لأن هذا التغيير يتغير بين الأشكال النشطة وغير النشطة، قد ترتبط أيضًا ببقية سطح البروتين، بروتينات أخرى موجودة قد تغير نشاط بروتينات عائلة راس، مثال واحد هو GDI (مثبط انفصال الناتج المحلي الإجمالي) ؛ هذه الوظيفة عن طريق إبطاء تبادل الناتج المحلي الإجمالي ل GTP وبالتالي، طالة أمد حالة عدم نشاط أفراد عائلة راس. قد توجد بروتينات أخرى تزيد من هذه الدورة.
ارتباطها على الغشاء
يرتبط بروتين راس في الغشاء الخلوي نتيجة لعملية إضافة البرينيل أو البرنلة (بالانجليزية:Prenylation) وعملية إضافة البالميتول (بالانجليزية: Palmitoylation) ل (HRAS و NRAS) ، أو نتيجة لمزيج من عملية البرنلة (بالانجليزية:Prenylation) وتسلسل متعدد القواعد مجاور لمكان عملية البرنلة (بالانجليزية:Prenylation) ل (KRAS). النهاية الطرفية سي ( C-terminal) CaaX box لبروتين راس في البداية يتم التعديل عليها من خلال عملية إضافة farnesyl group لجزيء الحمض الاميني سيستين (cysteine) في السايتوسول (cytosol)، مما يسمح لبروتين راس بالدخول الي غشاء الشبكة الاندوبلازمية و الاغشية الخلوية الاخرى بسهولة. ثم يتم فصل ثلاثي الببتيد (aaX) عن النهاية الطرفية سي (C-terminal) من خلال استخدام انزيمات خاصة لقطع البروتين ثم يتم إضافة مجموعة ميثل الي النهاية الطرفية سي (C-terminal) الجديدة من خلال الميثيل ترانسفيريز (بالانجليزية: methyltransferase). في هذه المرحلة يتم الانتهاء من معالجة KRas. التفاعلات الحركية الكهربائية بين الشحنات الموجبة للقواعد المسلسلة والشحنات السالبة في الجزء الداخلي من الغشاء البلازمي مسؤولة عن بقائها مستقرة على سطح الخلية بشكل ثابت . يتم التعديل ايضا على NRAS و HRAS على سطح جهاز جولجي من خلال عملية البالميتوليشن (بالانجليزية: Palmitoylation) لواحد أو اثنين من جزيئات الحمض الاميني سيستين (cysteine). وبذلك تصبح البروتينات مثبتة على الغشاء ويتم نقلها الي الغشاء البلازمي عن طريق حويصلات في المسار الافرازي. عملية الديبالميتوليشن ( بالانجليزية: Depalmitoylation) عن طريق acyl-protein thioesterases تسمح باطلاق سراح البروتين من الغشاء من اجل السماح له بالدخول الي دورة اخرى من عمليتي البالميتوليشن والديبالميتوليشن.[16] وهذه الدورة المستمرة تمنع تسرب NRAS وHRAS للاغشية الاخرى وتحافظ على استقرارها على سطح جهاز جولجي ، المسار الافرازي و الغشاء الخلوي
أفراد عائلة راس
ابرز اعضاء هذه الفصيلة التي تم ملاحظتها سريرياK RAS و NRAS و HRAS نتيجة لمشاركتها ودورها في انواع عديدة من السرطان.[17] إضافة إلى ذلك هناك افراد اخرى من هذه الفصيلة:[18]DIRAS1; DIRAS2; DIRAS3; ERAS; GEM; MRAS; NKIRAS1; NKIRAS2; NRAS; RALA; RALB; RAP1A; RAP1B; RAP2A; RAP2B; RAP2C; RASD1; RASD2; RASL10A; RASL10B; RASL11A; RASL11B; RASL12; REM1; REM2; RERG; RERGL; RRAD; RRAS; RRAS2
بروتينات راس والسرطان
حدوث الطفرات في عائلة راس من الجينات المسرطنة والتي تضم ( H-RAS وN-RAS و K-RAS ) عملية شائعة جدا، وجدت هذه الطفرات في 20% الي 30% من مجموع الاورام التي تصيب الانسان.[17] نتيجة لذلك فمن الطبيعي التوقع بان العلاج الدوائي الذي يحد من نشاط بروتين راس يمثل طريقة لتثبيط بعض انواع السرطان في الانسان. الطفرات النقطية الناتجة عن خلل في نيكليوتيد واحد في جينات راس هو الحالة الغير طبيعية الأكثر شيوعا في الجينات المسرطنة عند الانسان. مثبط بروتين راس trans- farnesylthiosalicylic acid (FTS, Salirasib)) اظهر اثر كبير في كمضاد لتطور الورم في الكثير من الخلايا السرطانية.[19][20]
التنشيط الغير مناسب
لقد اظهر التنشيط الزائد والغير مناسب لجينات راس دورا رئيسيا في نقل الاشارات الخاطئة التي تؤدي بدورها إلى نمو الخلايا وانقسامها بشكل كبير وهذا النمو الغير طبيعي ينتج عنه تكون الورم السرطاني.[21] الطفرات التي تحدث في عدد من الجينات المختلفة كما الحال في الجينات المترجمة لبروتين راس يمكن ان يكون لها هذا الاثر.الجينات السرطانية (Oncogenes) مثل p210BCR-ABL أو مستقبلات النمو erbB عبارة عن اشارات تنشيط قبلية ل راس. لذلك فانه إذا كانت منشطة اساسا فإن اشاراتها سوف تنتقل من خلال راس. يقوم الجين المثبط للورم NF1 بتشفير Ras-GAP حيث ان الطفرة في الورم الليفي العصبي ( neurofibromatosis) تعني ان راس اقل عرضة للتثبيط وعدم التفعيل. ممكن ان يحدث تضخم راس، لكن هذه الحالة تحدث فقط في بعض الاورام.
اخيرا، يمكن ان يتم تنشيط الجينات السرطانية راس من خلال الطفرات النقطية بحيث ان تفاعلات GTPase لا يمكن تحفيزها من خلال GAP وهذا يزيد العمر النصفي ل Ras-GTP النشط والذي حدثت فيه الطفرة.[22]
راس المنشط اساسا
وهو الذي يحتوي على على طفرات تمنع تحلل GTP وبالتالي تجعل راس في حالة تنشيط وتشغيل دائمة.
الطفرات الأكثر شيوعا تتواجد على جزيء G12 في P-loop وعلى جزيء الحفاز Q61.
- طفرة تحول الجلايسين (glycine) الي فالين (valine)على النهاية الطرفية 12 يحول مجال GTPase في Ras الغير حساس إلى غير فعال من خلال GAP مما يجعلها عالقة في حالة تنشيط دائمة. يحتاج بروتين راس إلى GAP للتثبيط لانه عبارة عن حفاز ضعيف نسبيا بمفرده، على عكس البروتينات الاخرى التي تحتوي على G-domain مثل الوحدة الفرعية الفا ل heterotrimeric G proteins.
- النهاية الطرفية 61[23] هي المسؤولة عن حفظ توازن المرحلة الانتقالية لتحلل GAP. لانه عموما التحفيز عن طريق الانزيم يتحقق من خلال تقليل الطاقة بين المواد المتفاعلة والنواتج، فان حدوث طفرة في Q61 إلى K يؤدي بالضرورة الي خفض مستوى تحلل Ras GTP الي مستويات لا معنى لها من الناحية الفسيولوجية.
علاجات السرطان التي تستهدف بروتين راس
لقد لوحظ ان الريوفايروس (Reovirus ) قد يكون علاجا فعالا للسرطان عندما اوضحت الدراسات انه يتكاثر بشكل جيد في بعض الخلايا السرطانية. يتكاثر بشكل خاص في الخلايا التي تحتوي على مسار راس نشط (وهو مسار خلوي يدخل في نمو الخلية وتمايزها).[24] يتكاثر الريوفايروس (Reovirus ) ويقتل الخلايا السرطانية التي تحتوي على بروتين راس نشط، عندما تموت الخلايا فاين بقايا الفيروس لها الحرية والإمكانية بالدخول الي خلايا سرطانية اخرى. يعتقد ان هذه الدورة من الإصابة والتكاثر و موت الخلايا تتكرر حتى يتم تدمير جميع الخلايا السرطانية التي يتواجد فيها مسار نشط من بروتين راس.
هناك فايروس اخر يستهدف الخلايا السرطانية التي تحتوي على مسار راس نشط ويقوم بتحليل الورم، وهو النوع الثاني من فيروس الهربس البسيط (HSV-2). الطفرات النشطة في بروتينات راس تلعب دورا في حوالي ثلثي انواع السرطانات التي تصيب الانسان وتتضمن السرطانات القابلة للانتشار. ريوليوسين ( Reolysin ) وهو نوع من انواع الريوفايروس و FusOn-H2 المستخدم حاليا في التجارب السريرية أو قيد التطوير لعلاج انواع مختلفة من السرطان.[25] بالإضافة إلى ذلك العلاج القائم على siRNA مضاد الطفرات في K-RAS (G12D) المسمى siG12D LODER حاليا هو قيد التجارب لعلاج سرطان البنكرياس متقدم المراحل.[26]
مراجع
- الطبعة العربية لدورية nature الشهرية العالمية للعلوم- عائلة راس نسخة محفوظة 12 يوليو 2019 على موقع واي باك مشين.
- Goodsell, David S. (1999-06-01). "The Molecular Perspective: The ras Oncogene". The Oncologist (باللغة الإنجليزية). 4 (3): 263–264. ISSN 1083-7159. PMID 10394594. مؤرشف من الأصل في 10 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Downward, Julian (2003/01). "Targeting RAS signalling pathways in cancer therapy". Nature Reviews Cancer (باللغة الإنجليزية). 3 (1): 11–22. doi:10.1038/nrc969. ISSN 1474-1768. مؤرشف من الأصل في 10 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Malumbres, Marcos; Barbacid, Mariano (2003/06). "Timeline: RAS oncogenes: the first 30 years". Nature Reviews Cancer (باللغة الإنجليزية). 3 (6): 459–465. doi:10.1038/nrc1097. ISSN 1474-1768. مؤرشف من الأصل في 10 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Chang, E. H.; Gonda, M. A.; Ellis, R. W.; Scolnick, E. M.; Lowy, D. R. (1982-08-01). "Human genome contains four genes homologous to transforming genes of Harvey and Kirsten murine sarcoma viruses". Proceedings of the National Academy of Sciences (باللغة الإنجليزية). 79 (16): 4848–4852. doi:10.1073/pnas.79.16.4848. ISSN 0027-8424. PMID 6289320. مؤرشف من الأصل في 13 يونيو 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - HARVEY, J. J. (1964/12). "An Unidentified Virus which causes the Rapid Production of Tumours in Mice". Nature (باللغة الإنجليزية). 204 (4963): 1104–1105. doi:10.1038/2041104b0. ISSN 1476-4687. مؤرشف من الأصل في 10 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Kirsten, W. H.; Schauf, V.; McCoy, J. (1970). "Properties of a murine sarcoma virus". Bibliotheca Haematologica (36): 246–249. ISSN 0067-7957. PMID 5538357. مؤرشف من الأصل في 23 يوليو 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cooper, G. M. (1982-08-27). "Cellular transforming genes". Science (باللغة الإنجليزية). 217 (4562): 801–806. doi:10.1126/science.6285471. ISSN 0036-8075. PMID 6285471. مؤرشف من الأصل في 02 يونيو 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Santos, Eugenio; Tronick, Steven R.; Aaronson, Stuart A.; Pulciani, Simonetta; Barbacid, Mariano (1982/07). "T24 human bladder carcinoma oncogene is an activated form of the normal human homologue of BALB- and Harvey-MSV transforming genes". Nature (باللغة الإنجليزية). 298 (5872): 343–347. doi:10.1038/298343a0. ISSN 1476-4687. مؤرشف من الأصل في 10 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Parada, Luis F.; Tabin, Clifford J.; Shih, Chiaho; Weinberg, Robert A. (1982/06). "Human EJ bladder carcinoma oncogene is homologue of Harvey sarcoma virus ras gene". Nature (باللغة الإنجليزية). 297 (5866): 474–478. doi:10.1038/297474a0. ISSN 1476-4687. مؤرشف من الأصل في 10 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Taparowsky, Elizabeth; Suard, Yolande; Fasano, Ottavio; Shimizu, Kenji; Goldfarb, Mitchell; Wigler, Michael (1982/12). "Activation of the T24 bladder carcinoma transforming gene is linked to a single amino acid change". Nature (باللغة الإنجليزية). 300 (5894): 762–765. doi:10.1038/300762a0. ISSN 1476-4687. مؤرشف من الأصل في 10 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Marshall, C. J.; Hall, A.; Weiss, R. A. (1982/09). "A transforming gene present in human sarcoma cell lines". Nature (باللغة الإنجليزية). 299 (5879): 171–173. doi:10.1038/299171a0. ISSN 1476-4687. مؤرشف من الأصل في 10 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Hall, Alan; Marshall, Christopher J.; Spurr, Nigel K.; Weiss, Robin A. (1983/06). "Identification of transforming gene in two human sarcoma cell lines as a new member of the ras gene family located on chromosome 1". Nature (باللغة الإنجليزية). 303 (5916): 396–400. doi:10.1038/303396a0. ISSN 1476-4687. مؤرشف من الأصل في 10 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Shimizu, K.; Goldfarb, M.; Perucho, M.; Wigler, M. (1983-01-01). "Isolation and preliminary characterization of the transforming gene of a human neuroblastoma cell line". Proceedings of the National Academy of Sciences. 80 (2): 383–387. doi:10.1073/pnas.80.2.383. مؤرشف من الأصل في 10 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Vetter, Ingrid R.; Wittinghofer, Alfred (2001-11-09). "The Guanine Nucleotide-Binding Switch in Three Dimensions". Science (باللغة الإنجليزية). 294 (5545): 1299–1304. doi:10.1126/science.1062023. ISSN 0036-8075. PMID 11701921. مؤرشف من الأصل في 30 مارس 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Spatio-temporal segregation of Ras signals: one ship, three anchors, many harbors". Current Opinion in Cell Biology (باللغة الإنجليزية). 18 (4): 351–357. 2006-08-01. doi:10.1016/j.ceb.2006.06.007. ISSN 0955-0674. مؤرشف من الأصل في 28 مارس 2013. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bos, J. L. (1989-09-01). "ras oncogenes in human cancer: a review". Cancer Research. 49 (17): 4682–4689. ISSN 0008-5472. PMID 2547513. مؤرشف من الأصل في 30 يوليو 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wennerberg, Krister; Rossman, Kent L.; Der, Channing J. (2005-03-01). "The Ras superfamily at a glance". J Cell Sci (باللغة الإنجليزية). 118 (5): 843–846. doi:10.1242/jcs.01660. ISSN 0021-9533. PMID 15731001. مؤرشف من الأصل في 22 يوليو 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Rotblat, Barak; Ehrlich, Marcello; Haklai, Roni; Kloog, Yoel (2008). "The Ras inhibitor farnesylthiosalicylic acid (Salirasib) disrupts the spatiotemporal localization of active Ras: a potential treatment for cancer". Methods in Enzymology. 439: 467–489. doi:10.1016/S0076-6879(07)00432-6. ISSN 0076-6879. PMID 18374183. مؤرشف من الأصل في 23 يوليو 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Blum, Roy; Jacob-Hirsch, Jasmine; Amariglio, Ninette; Rechavi, Gideon; Kloog, Yoel (2005-02-01). "Ras inhibition in glioblastoma down-regulates hypoxia-inducible factor-1alpha, causing glycolysis shutdown and cell death". Cancer Research. 65 (3): 999–1006. ISSN 0008-5472. PMID 15705901. مؤرشف من الأصل في 23 يوليو 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Molecular cell biology (الطبعة 4th ed). New York: W.H. Freeman. 2000. ISBN 071673706X. OCLC 41266312. مؤرشف من الأصل في 10 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة)صيانة CS1: نص إضافي (link) - Reuter, C. W.; Morgan, M. A.; Bergmann, L. (2000-09-01). "Targeting the Ras signaling pathway: a rational, mechanism-based treatment for hematologic malignancies?". Blood. 96 (5): 1655–1669. ISSN 0006-4971. PMID 10961860. مؤرشف من الأصل في 23 يوليو 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Omim - Neuroblastoma Ras Viral Oncogene Homolog; Nras [وصلة مكسورة] نسخة محفوظة 6 مارس 2019 على موقع واي باك مشين.
- Lal, Rohit; Harris, Dean; Postel-Vinay, Sophie; de Bono, Johann (October 2009). "Reovirus: Rationale and clinical trial update". Current Opinion in Molecular Therapeutics. 11 (5): 532–539. ISSN 2040-3445. PMID 19806501. مؤرشف من الأصل في 20 مايو 2016. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Thirukkumaran, Chandini; Morris, Don G. (2009). Gene Therapy of Cancer. (باللغة الإنجليزية). Humana Press. صفحات 607–634. doi:10.1007/978-1-59745-561-9_31. ISBN 9781934115855. مؤرشف من الأصل في 14 يونيو 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Search of: silenseed - List Results - ClinicalTrials.gov" (باللغة الإنجليزية). مؤرشف من الأصل في 09 أبريل 2018. اطلع عليه بتاريخ 08 أبريل 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ أرشيف=(مساعدة)
 بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة طب
بوابة طب بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي
