بكتيريا ممغنطة
البكتيريا الممغنطة (أو MTB ) هي مجموعة متعددة البكتيريا من ضمن البكتيريا التي توجه نفسها على طول خطوط المجال المغناطيسي و للمجال المغناطيسي للأرض . [1] اكتشفت في عام 1963من قبل سالفاتوري بيليني و تم أعادة اكتشافها من قبل ريتشارد بلاكمور في عام 1975. ويعتقد أن هذه المحاذاة تساعد هذا النوع من الكائنات في الوصول إلى مناطق تركيز الأكسجين بشكل امثل. [2] و لأداء هذه المهمة تحتوي هذه البكتيريا على نوع من عضيات تسمى المغنطيسية و التي تحتوي على بلورات مغناطيسية. فالظاهرة البيولوجية للكائنات الحية الدقيقة التي تميل إلى التحرك وذلك استجابة للخصائص المغناطيسية للبيئة باسم المغنطيسية . و على الرغم من ذلك فإن هذا المصطلح يعتبر مضلل من حيث أن كل تطبيق آخر لمصطلح سيارات الأجرة ينطوي على آلية استجابة التحفيز. على عكس استقبال المغناطيسات للحيوانات تحتوي البكتيريا على مغناطيس ثابت فمن خلال المغناطيس يجبر البكتيريا على المحاذاة حتى يتم سحب الخلايا الميتة إلى المحاذاة ، تمامًا مثل كإبرة البوصلة. [3]
المقدمة
ففي عام 1963 تم وضع أول وصف للبكتيريا الممغنطة بواسطة سالفاتور بيليني من جامعة بافيا . [4] [5] وذلك أثناء مراقبة رواسب المستنقعات تحت المجهر، فمن خلال ذلك لاحظ بيليني مجموعة من البكتيريا التي من الواضح أنها توجه نفسها في اتجاه فريد. فمن خلال ما شاهدة أدرك أن هذه الكائنات الدقيقة تتحرك وفقًا لاتجاه القطب الشمالي . وبعد ذلك أطلق عليها "البكتيريا الحساسة للمغناطيسية". فقد كانت المنشورات أكاديمية (تمت مراجعتها من قبل الأقران من قبل لجنة التحرير في Istituto di Microbiologia تحت مسؤولية مدير المعهد الأستاذ بيانكي ، كالمعتاد في الجامعات الأوروبية في ذلك الوقت) وتم نقلها من اللغة الإيطالية إلى ملخصات قصيرة باللغة الإنجليزية والفرنسية والألمانية وذلك في الجريدة الرسمية لمؤسسة معروفة . ولكن يبدو أنه لم تجذب سوى القليل من الاهتمام إلى أن تم توجيه انتباه ريتشارد فرانكل إليها وذلك في عام 2007. قام فرانكل بترجمتها إلى الإنجليزية ونشرت الترجمات في المجلة الصينية لعلوم المحيطات وعلم Limnology . [6] [7] [8] [9]
ولاحظ ريتشارد بلاكيمور و الذي كان في ذلك الوقت خريجًا في علم الأحياء الدقيقة [10] في جامعة ماساتشوستس في أمهيرست ، والذي يعمل في مؤسسة وودز هول لعلوم المحيطات التي تتوفر فيها مجموعة المنشورات ذات الصلة والتي ينشرها معهد علم الأحياء الدقيقة بجامعة بافيا موجودة. لاحظت وجود كائنات حية دقيقة تسير في اتجاه المجال المغناطيسي للأرض. لم يذكر Blakemore بحث Bellini في تقريره الخاص الذي نشره في مجلة Science ، ولكنه تمكن من مراقبة سلاسل المغنطيسية وذلك باستخدام المجهر الإلكتروني. [8] [11] شروط بيليني لهذا السلوك ، وبالتحديد (بالإيطالية: batteri magnetosensibili) ، (بالفرنسية: bactéries magnétosensibles) أو (بالفرنسية: bactéries aimantées) ، German English: (منشور بيليني الأول ، الصفحة الأخيرة) ، تم نسيانه ، وتم تبني "مغناطيسية" Blakemore من قبل المجتمع العلمي. وقد خضعت هذا النوع من البكتيريا لتجارب عديدة .بل إنهم كانوا على متن مكوك الفضاء لفحص خصائصها المغناطيسية في غياب الجاذبية ، وعلى الرغم من ذلك لم يتم التوصل إلى نتيجة نهائية. [12]
فحساسية البكتيريا الممغنطة تجاه المجال المغناطيسي للأرض تحدث من حقيقة أن هذا النوع من البكتيريا تعجل سلاسل بلورات المعادن المغناطيسي في داخل خلاياها. وتفيد التقارير حتى الآن بأن جميع البكتيريا الممغنطة تترسب إما المغنتيت أو الجريجيت .كما يمكن الاحتفاظ على هذه البلورات وفي بعض الأحيان سلاسل البلورات ، في السجل الجيولوجي كأحافير مغناطيسية . أقدم المغنطيسية magnetofossils الواضحة تعود إلى العصر الطباشيري الأسرة الطباشير في جنوب إنكلترا، [13] وان كانت هناك تقارير اقل تأكيدا عن وجود أحافير أ المغنطيسية magnetofossils تعود إلى 1.9 مليار سنة جونفلينت تشرت . [14] كانت هناك أيضًا ادعاءات بوجودها على المريخ استنادًا إلى شكل جزيئات المغنتيت داخل نيزك المريخ ALH84001 ، ولكن هذه الادعاءات في موضع جدل كبير. [15]
مادة الاحياء
توجد العديد من الأشكال المختلفة لبروميد الميثيل والتي قد تختلف من حيث العدد والشكل ونمط الجسيمات المغناطيسية البكتيرية التي تحتوي عليها [16] ويمكن تقسيم المركبات ثلاثي الفينيل متعدد الكلور إلى فئتين .وطبقا اذلك فان كانت تنتج جزيئات من المغنتيت ( Fe3O4 </br> Fe3O4 </br> Fe3O4 ) أو الجريجيت ( Fe3S4 </br> Fe3S4 </br> Fe3S4 ) على الرغم من بعض الأنواع قادرون على إنتاج كليهما. يمتلك المغناطيس لحظة مغناطيسية بثلاثة أضعاف حجم الجريجيت. [15]
في العادة توجد البكتيريا الممغنطة إي المنتجة للمغنطيسية في منطقة انتقالية أوضاع مؤكسدة-نقص الأكسجين (OATZ)، وهى المنطقة الانتقالية بين المياه أو الرواسب الغنية بالأكسجين والمتعطشة للأكسجين. ولا يتمكن الكثير من بروميد الميثيل من قدرة على البقاء إلا في البيئات ذات الأكسجين المحدود جدا ، ولا يوجد بعضها إلا في البيئات اللاهوائية تمامًا. تم أفترض أن الميزة التطورية لامتلاك نظام من من الجسيمات المغنطيسية ترتبط بالقدرة على التنقل بكفاءة داخل هذه المنطقة من التدرجات الكيميائية الحادة من خلال تبسيط البحث الثلاثي الأبعاد المحتمل لظروف أكثر ملاءمة لبعد واحد. (انظر § Magnetism لوصف هذه الآلية. ) كما يمكن لبعض أنواع البكتيريا المغنطيسية أن تنتج مغناطيسا حتى في الظروف اللاهوائية ، وذلك باستخدام أكسيد النيتريك أو النترات أو الكبريتات. كمستقبل نهائي للإلكترونات . أن مركبات القصدير الثلاثي البيوتيل التي تمعدن الغريغات تكون عادة لاهوائية بشكل صارم. [17]
وقد تم الأشاره على أن هذه المادة قد تطورت في مطلع عصور الأرخبيل Archean . كما أن الزيادة في الأكسجين الجوي تعني أن للكائنات الحية ميزة تطورية في امتلاكها القدرة المغناطيسية. [18] تطورت الجسيمات المغناطيسية أولا كآلية دفاعية استجابة للزيادة في أنواع الأكسجين التفاعلية (ROS) التي نتجت عن حدث الأكسجين العظيم. [19] و بدأت الكائنات الحية تخزين الحديد في شكل ما ، وتم تكييف هذا الحديد داخل الخلايا في ما بعد ليشكل مغناطيس. قد تكون هذه MTB المبكرة قد شاركت في تكوين أول خلايا حقيقية النواة. [14] تم العثور على المغناطيس الحيوي المنشأ مماثل لتلك الموجودة في البكتيريا الممغنطة في الكائنات الحية الأعلى ، من الطحالب السرطانية إلى التراوت . [20] كما ان التقارير في البشر والحمام أقل تقدمًا بكثير. [21]
قد تقوم البكتيريا الممغنطة بتنظيم جزيئاتها المغناطيسية عن طريق سلاسل خطية. ولذلك فإن عزم ثنائي القطب المغناطيسي لفي اللية هو مجموع عزم ثنائي القطب لكل BMP ،وهذا الشيء كفيل لتوجيه الخلية بشكل سلبي والتغلب على القوى الحرارية العرضية الموجودة في البيئة المائية. [17] أثناء وجود أكثر من سلسلة ستدفع قوى التنافر بين سلاسل هذه الهياكل إلى حافة الخلية ، مسببة التورم . [15]
توجد جميع الجينات وشيجة الصلة بالمغناطيسية في MTB في منطقة 80 كيلوباس تقريبًا في الجينوم مسمى بجزيرة المغناطيس . [22] هناك ثلاثة عوامل رئيسية في الجزيرة الصغرى المغناطيسية: الأوبون mamAB ، الأوبون mamGFDC ، الأوبرا mms6. كذلك هناك 9 جينات ضرورية لتكوين ووظيفة المغنطومات الحديثة: mamA و mamB و mamE و mamI و mamK و mamM و mamO و mamP و mamQ. [23] بالإضافة إلى هذه الجينات التسعة المحفوظة بشكل جيدً في جميع MTB ، و هناك أكثر من 30 جينًا إجماليًا قد ساهمت في حدوث الخلايا المغنطيسية في MTB. وتمثل هذه الجينات غير الأساسية مسؤولة عن الاختلاف في حجم وشكل بلورات المغنتيت / الجريجيت ، بالإضافة إلى المحاذاة المحددة للمغناطيسية في الخلية.
ويظهر تنوع بروميد الميثيل من خلال عدد كبير من الأنماط المورفوتية المختلفة الموجودة في العينات البيئية للمياه أو في الرواسب. وتشمل هذه الأنماط المورفولوجية الشائعة التي يتم ملاحظتها عموما الخلايا الكروية أو البيضاوية ( العصعص ) ذات الأبعاد المختلفة ، على شكل قضيب ( عصيات ) ، والبكتيريا اللولبية . أحد الأنواع المورفولوجية الأكثر تميزا هي بكتيريا متعددة الخلايا على ما يبدو يشار إليها باسم بدائية النوى المغناطيسية متعددة الخلايا (MMP).
بغض النظر عن مورفولوجيا هذه السلالة ، فإن جميع السلالات التي خضعت للدراسة حتى الآن متحركة عن طريق السوط وهي بكتيريا سالبة الجرام من السلالات المختلفة. وعلى الرغم من أن معظم الأنواع المعروفة تعتبر بكتيريا بروتينية ، على سبيل المثال Magnetospirillum Magneticum a alphaproteobacterium ،قد يمتلك أعضاء من سلالات مختلفة مجموعة جينات magnetosome ، مثل Candidatus Magnetobacterium bavaricum a Nitrospira. [24] و يختلف ترتيب السوط وقد يمكن ان يكون قطبيًا أو ثنائي القطب أو في خصل . وفي عام 1991 تم إجراء أول تحليل للتطور الوراثي على البكتيريا الممغنطة باستخدام مقارنات تسلسل الجينات 16R rRNA بواسطة P. Eden et al
تعتبر ترتيب الجسيمات المغناطيسية داخل الخلية البكتيرية هيه سمة أخرى قد تظهر تنوعا كبيرا . في غالبية MTB ، يتم محاذاة المغنسيوم في سلاسل ذات أطوال وأرقام مختلفة على طول المحور الطويل للخلية ، وهو يعتبر الاتجاه الأكثر فعالا بالمغناطيس. وعلى الرغم من كل ذلك ، تحدث المجاميع أو مجموعات المغنطيسية المتناثرة في بعض MTB ،عادتا في جانب واحد في الخلية ، والذي في الغالب يتوافق مع موقع إدخال السوط. إلى جانب المغنسيوم ، فإن الأجسام الكبيرة التي تحتوي على عنصر الكبريت ، أو بولي فوسفات ، أو بولي هيدروكسي بوتيرات شائعة الوجود في MTB.
الخلايا الكوكبية التي تمتلك حزمتين من السوط على جانب مسطح إلى حد ما قد تعتبر النوع الأكثر وفرة وشيوع من MTB التي تحدث في العينات البيئية ، وخاصة الرواسب. وقد أدى هذا النوع "الجلدي" إلى تكاثر جنس "المكورات الميكروية" لهذه البكتيريا. في المقابل ، اثنان من MTB أكثر وضوحًا من الناحية الشكلية ، يتم ملاحظتهما بانتظام في العينات الطبيعية ، ولكن لا يتم عزلهما أبدًا في الثقافة النقية ، وهما MMP وقضيب كبير يحتوي على كميات وفيرة من المغنطومات على شكل خطاف ( Magnetobacterium bavaricum ).
المغناطيسية
ويخضع التطور الجسدي للبلورة المغناطيسية لعاملين: الأول نحو محاذاة القوة المغناطيسية للجزيئات بالاقتران مع البلورة النامية ، في حين يقلل الآخر القوة المغناطيسية للبلورة ، مما قد يسمح بارتباط الجزيء بينما يواجه قوة مغناطيسية معاكسة . في الطبيعة ، و هذا في طبيعته ستسبب في وجود مجال مغناطيسي يحيط بمحيط المجال بسماكة تقارب 150 نانومتر من المغنتيت ، حيث تغير الجزيئات اتجاهاتها بشكل تدريجى.و لهذا السبب لا يكون الحديد مغناطيسيًا في غياب المجال تطبيقي. وعلى النحو المماثل لا تظهر الجسيمات المغناطيسية الصغيرة للغاية لا تظهر عليها علامات مغنطة في درجة حرارة الغرفة ؛ تغيير قوتها المغناطيسية باستمرار بالحركات الحرارية الكامنة في تكوينها. [15] بدلاً من ذلك ، فإن البلورات المغنطيسية فيMTB يتراوح حجمها بين 35 و 120 نانومتر ،أي أنها كبيرة بما يكفي لان يكون لها مجال مغناطيسي وفي الوقت نفسه صغير بما يكفي لتظل مجالًا مغناطيسيًا واحدًا . [17]
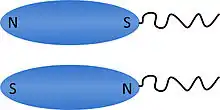
وميلان المجال المغنطيسي للأرض في نصفي الكرة الأرضية كليهما يختار واحد من اثنين من أقطاب المحتملة للخلايا magnetotactic (فيما يتعلق القطب سوطي للخلية)، توجيه تمعدن حيوي من magnetosomes .
المحور الهوائي هو الاستجابة التي تهاجر بها البكتيريا إلى تركيز أمثل للأوكسجين في تدرج الأكسجين. و أظهرت تجارب فايروسي بوضوح أن التقسيمات المغنطيسية والمحور الهوائي يعملان بالاقتران مع البكتيريا المغناطيسية . وقد تبين أنه يمكن للبكتيريا المغناطيسية للسباحة ذات الاتجاه الواحد التي تسبح في قطرات الماء يمكنها أن تعكس اتجاه سباحتها وتسبح إلى الوراء تحت ظروف الاختزال (أقل من تركيز الأكسجين الأمثل) ، على عكس الظروف الثورية (أكبر من تركيز الأكسجين الأمثل). يشار إلى السلوك الذي لوحظ في هذه السلالات البكتيرية باسم المغناطيسية الهوائية
توجد أليتين مغناطيسيتان هوائيتان مختلفتان - وقد تعرف باسم القطبية والمحورية - في سلالات MTB المختلفة. [25] بعض السلالات التي تسبح باستمرار في اتجاه واحد على طول المجال المغناطيسي (إما باتجاه الشمال [NS] واتجاه الجنوب [SS]) - بصفة رئيسية. المكورات المغناطيسية - هذه البكتيريا المغناطيسية ستنتقل ستتحرك على طول خطوط في المجال المغناطيسي للأرض وفقًا لتوجهاتها ، لكنها ستنحرف كمجموعة وتنعكس اتجاهها إذا تعرضت لمجال مغناطيسي محلي أكثر قوة وموجه عكس الاتجاه . وبهذه الطريقة ،تستمر في التحرك في نفس الاتجاه المغناطيسي ، ولكن نسبيًا بدلاً من ذلك إلى المجال المحلي . وهذه المركبات التي تسبح في أي من الاتجاهين على طول خطوط المجال المغناطيسي مع انعكاس متكرر وعفوي لاتجاه السباحة دون العودة - على سبيل المثال ، spirilla المياه العذبة - تكتيك هوائي مغنطيسي محوري ولا ينطبق التمييز بين NS و SS على هذه البكتيريا. في حين يوفر المجال المغنطيسي محورا واتجاها لحركة البكتيريا القطبية المغنطيسية الهوائية. في حين يوفر محورا للحركة فقط لأنواع البكتيريا المحورية و في كلتا الحالتين ، يزيد التقسيط المغنطيسي من كفاءة المحور الهوائي في تدرجات التركيز الرأسي عن طريق تقليل البحث ثلاثي الأبعاد إلى بُعد واحد.
اقترح العلماء أيضا يوسع النموذج الموصوف لمحور ماكنيتو-أيروتكسيس إلى مركبات أكثر تعقيدا. وفي هذه الحالة ،فإن حركة أحادية الاتجاه لبروميد الميثيل في نقطة الماء لن تكون سواء جانبًا واحدًا من الاستجابة المتطورة للتحكم في بالاختزال. ومن بين التلميحات إلى الوظيفة المحتملة للتقسيسات المغناطيسية القطبية أن معظم الكائنات الحية الدقيقة المجهرية تتسم بامتلاكها إما شوائب كبريتية كبيرة أو مغناطيسات تتكون من كبريتات الحديد. لذلك ، قد يتم التكهن بأن عملية التمثيل الغذائي لهذه البكتيريا ، سواء كانت كيميائية غير متجانسة أو تغذوية ، تعتمد بشدة على امتصاص مركبات الكبريت المخفضة ، والتي تحدث في العديد من الموائل فقط في المناطق الأعمق عند أو تحت OATZ بسبب الأكسدة الكيميائية السريعة لـ هذه الأنواع الكيميائية المخفضة بواسطة الأكسجين أو المؤكسدات الأخرى في الطبقات العليا.
على سبيل المثال ، تستخدم الكائنات الحية الدقيقة التي تنتمي إلى جنس Thioploca النترات والتي يتم تخزينها بداخل الخلايا ، لأكسدة الكبريتيد ، وقد طورت غمدًا رأسيًا توجد فيه حزم من الخيوط المتحركة. من المفترض أن Thioploca يستخدم هذه الأغماد للتحرك بكفاءة في اتجاه عمودي في الرواسب ، وبالتالي تراكم الكبريتيد في الطبقات العميقة والنترات في الطبقات العليا. بالنسبة لبعض MTB ، قد يكون من الضروري أيضًا القيام برحلات إلى مناطق غير سامة من موطنها لتراكم مركبات كبريت مخفضة.
مراجع
- Lin, Wei; Zhang, Wensi; Zhao, Xiang; Roberts, Andrew; Paterson, Greig; Bazylinski, Dennis; Pan, Yongxin (March 2018). "Genomic expansion of magnetotactic bacteria reveals an early common origin of magnetotaxis with lineage-specific evolution". The ISME Journal. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Dusenbery, David B. (2009). Living at micro scale : the unexpected physics of being small. Cambridge, Mass.: Harvard University Press. ISBN 978-0-674-03116-6. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Dusenbery, David B. (1996).
- Bellini, S. (1963). Su di un particolare comportamento di batteri d'acqua dolce [On a unique behavior of freshwater bacteria] (PDF) (باللغة الإيطالية). Italy: Institute of Microbiology, University of Pavia. مؤرشف من الأصل (PDF) في 28 نوفمبر 2015. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bellini, S. (1963). Ulteriori studi sui "batteri magnetosensibili" [Further studies on magnetosensitive bacteria] (PDF) (باللغة الإيطالية). Italy: Institute of Microbiology, University of Pavia. مؤرشف من الأصل (PDF) في 28 نوفمبر 2015. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bellini, Salvatore (27 March 2009). "On a unique behavior of freshwater bacteria". Chinese Journal of Oceanology and Limnology. 27 (1): 3–5. Bibcode:2009ChJOL..27....3B. doi:10.1007/s00343-009-0003-5. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bellini, Salvatore (27 March 2009). "Further studies on "magnetosensitive bacteria"". Chinese Journal of Oceanology and Limnology. 27 (1): 6–12. Bibcode:2009ChJOL..27....6B. doi:10.1007/s00343-009-0006-2. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Frankel, Richard B. (27 March 2009). "The discovery of magnetotactic/magnetosensitive bacteria". Chinese Journal of Oceanology and Limnology. 27 (1): 1–2. Bibcode:2009ChJOL..27....1F. doi:10.1007/s00343-009-0001-7. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Komeili, Arash (January 2012). "Molecular mechanisms of compartmentalization and biomineralization in magnetotactic bacteria". FEMS Microbiology Reviews. 36 (1): 232–255. doi:10.1111/j.1574-6976.2011.00315.x. PMID 22092030. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Schaechter, Moselio, Ed.-in-Chief (2009). Encyclopedia of Microbiology, 3rd Ed (باللغة الإنجليزية). volume V. Academic Press, Elsevier. صفحة 230. مؤرشف من الأصل في 14 مايو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Blakemore, Richard (1975). "Magnetotactic Bacteria". Science. 190 (4212): 377–379. Bibcode:1975Sci...190..377B. doi:10.1126/science.170679. PMID 170679. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Urban (2000). "Adverse effects of microgravity on the magnetotactic bacterium Magnetospirillum magnetotacticum". Acta Astronautica. 47 (10): 775–780. Bibcode:2000AcAau..47..775U. doi:10.1016/S0094-5765(00)00120-X. PMID 11543576. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Kopp, R. E. & Kirschvink, J. L. (2007). "The identification and biogeochemical interpretation of fossil magnetotactic bacteria" (PDF). Earth-Science Reviews. 86 (1–4): 42–61. Bibcode:2008ESRv...86...42K. doi:10.1016/j.earscirev.2007.08.001. مؤرشف من الأصل (PDF) في 31 مايو 2012. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Chang, S. R. & J. L. Kirschvink (1989). "Magnetofossils, the magnetization of sediments, and the evolution of magnetite biomineralization". Annual Review of Earth and Planetary Sciences. 17: 169–195. Bibcode:1989AREPS..17..169C. doi:10.1146/annurev.ea.17.050189.001125. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cat Faber, Living Lodestones: Magnetotactic bacteria نسخة محفوظة 2006-05-07 على موقع واي باك مشين., Strange Horizons, 2001
- Schüler, Dirk (2002). "The biomineralization of magnetosomes in Magnetospirillum gryphiswaldense". Int. Microbiology. 5 (4): 209–214. doi:10.1007/s10123-002-0086-8. PMID 12497187. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bazylinski, Dennis (1995). "Controlled biomineralization of magnetic minerals by magnetotactic bacteria". Chemical Geology. 132 (1–4): 191–198. Bibcode:1996ChGeo.132..191B. doi:10.1016/S0009-2541(96)00055-1. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lin, Wei; Paterson, Greig; Zhu, Qiyun; Wang, Yinzhao; Kopylova, Evguenia; Li, Ying; Rob, Knight (February 2017). "Origin of microbial biomineralization and magnetotaxis during the Archean". Proceedings of the National Academy of Sciences. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Guo, Fang; Yang, Wei; Jiang, Wei; Geng, Shuang; Peng, Tao; Ji, Li (February 2012). "Magnetosomes eliminate intracellular reactive oxygen species in Magnetospirillum gryphiswaldense MSR-1". Environmental Microbiology. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Magnetic characterization of isolated candidate vertebrate magnetoreceptor cells". Proc Natl Acad Sci U S A. 109 (30): 12022–7. Jul 24, 2012. Bibcode:2012PNAS..10912022E. doi:10.1073/pnas.1205653109. PMID 22778440. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Clusters of iron-rich cells in the upper beak of pigeons are macrophages not magnetosensitive neurons". Nature. 484 (7394): 367–70. Apr 11, 2012. Bibcode:2012Natur.484..367T. doi:10.1038/nature11046. PMID 22495303. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Schübbe, Sabrina; Kube, Michael; Scheffel, André; Wawer, Cathrin; Heyen, Udo; Meyerdierks, Anke; Madkour, Mohamed H.; Mayer, Frank; Reinhardt, Richard (1 October 2003). "Characterization of a spontaneous nonmagnetic mutant of Magnetospirillum gryphiswaldense reveals a large deletion comprising a putative magnetosome island". Journal of Bacteriology. 185 (19): 5779–5790. doi:10.1128/JB.185.19.5779-5790.2003. PMID 13129949. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lefèvre, Christopher T.; Trubitsyn, Denis; Abreu, Fernanda; Kolinko, Sebastian; Jogler, Christian; de Almeida, Luiz Gonzaga Paula; de Vasconcelos, Ana Tereza R.; Kube, Michael; Reinhardt, Richard (May 2013). "Comparative genomic analysis of magnetotactic bacteria from the Deltaproteobacteria provides new insights into magnetite and greigite magnetosome genes required for magnetotaxis". Environmental Microbiology. 25 (10): 2712–2735. doi:10.1111/1462-2920.12128. PMID 23607663. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Jogler, C.; Wanner, G.; Kolinko, S.; Niebler, M.; Amann, R.; Petersen, N.; Kube, M.; Reinhardt, R.; Schuler, D. (2010). "Conservation of proteobacterial magnetosome genes and structures in an uncultivated member of the deep-branching Nitrospira phylum". Proceedings of the National Academy of Sciences. 108 (3): 1134–1139. Bibcode:2011PNAS..108.1134J. doi:10.1073/pnas.1012694108. PMID 21191098. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Frankel, R. B., D. A. Bazylinski, and D. Schüler (1998). "Biomineralization of magnetic iron minerals in magnetotactic bacteria". J. Supramolecular Science. 5 (3–4): 383–390. doi:10.1016/S0968-5677(98)00036-4. مؤرشف من الأصل في 15 مايو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link)
 صور وملفات صوتية من كومنز
صور وملفات صوتية من كومنز
.svg.png.webp) بوابة علم الأحياء الدقيقة
بوابة علم الأحياء الدقيقة
