العصر الطباشيري
الطباشيري أو الكريتاسي (باللاتينية: Cretaceous)، وهو الثالث والأخير من عصور حقبة الحياة الوسطى الثلاثة، امتد من ≈ 145 إلى 66 مليون سنة مضت، لمدة 79 مليون سنة تقريبا[6][7]. يسبقه الجوراسي، ويليه الباليوجيني من حقبة الحياة الحديثة.
| العصر الطباشيري | |||||||
|---|---|---|---|---|---|---|---|
| Cretaceous | |||||||
| الرمز | K | ||||||
| المستوى الزمني | عصر | ||||||
| الحقبة | الحياة الوسطى | ||||||
| -الدهر | دهر البشائر | ||||||
| علم الطبقات | |||||||
| البداية | ≈145 م.س.مضت | ||||||
| النهاية | |||||||
| المدة | 79 م.س تقريبا | ||||||
| مسند أول ظهور (FAD) |
Calpionella alpina[1] | ||||||
|
|||||||
|
|||||||
| أهم الصخور | الطبشور | ||||||
| الأقسام الفرعية |
| ||||||
| الجغرافيا القديمة والمناخ | |||||||
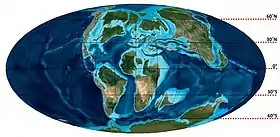 تشكل جغرافيا الأرض في نهاية الطباشيري (90 م.س مضت). تشكل جغرافيا الأرض في نهاية الطباشيري (90 م.س مضت). | |||||||
| نسبة الأكسجين في الغلاف الجوي |
تقريبا 30% حجما[2][3] (150 % من المستوى الحديث) | ||||||
| نسبة ثاني أكسيد الكربون في الغلاف الجوي |
تقريبا 1700 جزء في المليون[4] (أعلى 6 مرة من مستوى ما قبل الثورة الصناعية) | ||||||
| نسبة درجة حرارة سطح الأرض |
تقريبا 18 درجة مئوية[5] (4 درجات مئوية عن المستوى الحديث) | ||||||
| أحداث جيوديناميكية | |||||||
| |||||||
| الحيوانات و النباتات | |||||||
 العصر الطباشيري | |||||||
| التطور | |||||||
| |||||||
الأحداث الرئيسية في الطباشيري عرض • ناقش • -150 — – -140 — – -130 — – -120 — – -110 — – -100 — – -90 — – -80 — – -70 — – -60 — الأحداث الرئيسية في الطباشيري. مراحل وافقت عليها اللجنة الدولية للطبقات. مقياس المحور: منذ ملايين السنين. | |||||||
| (م.س : مليون سنة) | |||||||
كان العصر الطباشيري ذو مناخ دافئ نسبيا، وقد أدى إلى ارتفاع منسوب مياه البحار، الذي خلق عدد من البحار الداخلية الضحلة. وكانت هذه المحيطات والبحار مأهولة بزواحف بحرية تعتبر منقرضة في زمننا الحالي، مثل الأمونيتات والرودست، بينما الديناصورات لا تزال تواصل سيطرتها على اليابسة. خلال هذا الوقت، ظهرت مجموعات جديدة من الثدييات والطيور، وكذلك النباتات المزهرة.
انتهى العصر الطباشيري (مع نهاية حقبة الحياة الوسطى) بحدث انقراض العصر الطباشيري-الباليوجيني، الانقراض الجماعي الكبير الذي قضى على العديد من المجموعات، كالديناصورات الغير طيرية، والتيروصورات والزواحف البحرية الكبيرة. تم تعريف نهاية العصر الطباشيري بالحدود الطباشيرية الباليوجينية (K–Pg) المباغتة، التي تعتبر رمز جيولوجي مرتبط بالانقراض الجماعي الذي يقع بين حقبتي الحياة الوسطى ولحياة الحديثة.
التسمية
يعود مصطلح "الطباشيري" (Cretaceous) من اللاتينية (creta) التي تعني طباشير[8] وتختصر عادة بالحرف (K) بسبب الترجمة الألمانية (Kreide). وقد صاغه الجيولوجي البلجيكي جان دوماليوس هالوي في عام 1822،[9] وقد حدد العصر الطباشيري كعصر منفصلة لأول مرة، باستخدام الطبقات الموجودة في حوض باريس،[10] وبالتالي اطلق عليه اسم الطباشيري بسبب طبقات الطباشير الواسعة (كربونات الكالسيوم ترسب بسبب اصداف اللافقاريات البحرية، وخاصة البذور الجيرية [الإنجليزية])، التي وجدت في فترة الطباشيري العلوي في أوروبا الغربية.[lower-alpha 1]
الأقسام الفرعية
التقسيمات الفرعية لعصر الطباشيري من الأحدث إلى الأقدم هي:
| العصر | الفترة | المرحلة | أهم الأحداث | البداية (م.س.مضت) |
|---|---|---|---|---|
| الباليوجيني | الباليوسيني | الداني | أحدث | |
| الطباشيري | المتأخر | الماسترخي | أنتشرت كاسيات البذور (النباتات المزهرة)، إلى جانب بعض أنواع الحشرات الجديدة. ظهور الكثير من الأسماك العظمية الحديثة. شاع كل من الأمونيتات، والسهميات، وذوات الصدفتين، وقنافذ البحر، والإسفنجيات والرودست. تطورت أنواع جديدة من الديناصورات على الأرض (مثل التيرانوصوريات، وحجريات الترصع، والهادروصوريات ، وقرنيات الوجه)، وكذلك فعلت أسلاف التماسيح اليوسوكيات (التماسيح الحديثة)، وكذلك ظهرت في البحار الموزاصوريات وأسماك القرش الحديث. تعايشت الطيور المسننة والغير مسننة مع التيروصورات. ظهرت الثدييات الجرابية والمشيمة مثل الكظاميات ، والشقبانيات والوحشيات الحقيقية. تفككن قارة غندوانا العظمى. بداية تكون جبال لاراميد و تكون جبال السيفارية لجبال روكي في أمريكا الشمالية. أصبحت مستويات ثاني أكسيد الكربون في الغلاف الجويٍ قريبة من مستويات عصرنا الحالي. بدأت الديناصورات تنقرض بنهايات هذا العصر مع حدث انقراض العصر الطباشيري-الباليوجيني. | |
| الكامباني | 83.6 ± 0.2 | |||
| السانتوني | ||||
| الكونياكي | 89.8 ± 0.3 | |||
| التوروني | ||||
| السينوماني | ||||
| المبكر | الألبي | |||
| الأبتي | ≈125 | |||
| الباريمي | ≈129.4 | |||
| الهاتريفي | ≈132.9 | |||
| الفالانجيني | ≈139.8 | |||
| البرياسي | ≈145 | |||
| الجوراسي | المتأخر | التيثوني | أقدم | |
الجيولوجياء
علم الطبقات

كما هو الحال مع العصور الجيولوجية الأقدم، فقد تم تحديد الأسطح الصخرية للطباشيري بشكل جيد ولكن العمر الدقيق لقاعدة النظام غير مؤكد ببضعة ملايين السنين. ولا يوجد انقراض كبير أو انفجار تنوعي يفصل الطباشيري عن الجوراسي. ومع ذلك، فقد تم تعريف الجزء العلوي من النظام بشكل حاد، وتم وضعها في طبقة غنية بالإيريديوم الموجودة في جميع أنحاء العالم والتي يعتقد أنها مرتبطة بحفرة صدمة تشيكشولوب، مع حدودها التي تحد من أجزاء من شبه جزيرة يوكاتان وخليج المكسيك. وتم تأريخ هذه الطبقة عند 66.043 مليون سنة مضت.[11]
تم اقتراح العمر 140 مليون سنة مضت للحدود الجوراسية الطباشيرية بدلاً من 145 م.س مضت المقترح في عام 2014 بناءً على دراسة طبقية لتكوين فاكا مورتا [الإنجليزية] في حوض نيوكوين، الأرجنتين.[12] إن فيكتور راموس أحد مؤلفي الدراسة الذين يقترحون العمر بحدود 140 م.س مضت، معتبرين هذه الدراسة "خطوة أولى" نحو تغيير العمر رسميا في الاتحاد الدولي للعلوم الجيولوجية.[13]
تشكل الصخور


أدى ارتفاع مستوى سطح البحر والمناخ الدافئ في العصر الطباشيري إلى تغطية مناطق كبيرة من القارات بالبحار الضحلة الدافئة، التي وفرت موائل لكثير من الكائنات البحرية. وأحد أسباب تسمية العصر الطباشيري بهذا الاسم هو ترسبات الطباشير المنتشر خلال هذا العصر في أوروبا، ولكن كانت الرواسب في أجزاء كثيرة من العالم خلال هذا العصر من الحجر الجيري البحري، وهو صخر يتكون تحت الظروف البحرية الضحلة الدافئة. كانت هناك مساحات واسعة لمثل هذا الترسيب بسبب ارتفاع مستوى سطح البحر، وبسبب العمر الصغير والسمك الكبير لطبقة النظام، فإن الصخور الطباشيرية تظهر في العديد من المناطق حول العالم.
تعتبر صخور الطباشير نوع مميز (ليست حصرا) للعصر الطباشيري. التي تكونت من البذور الجيرية [الإنجليزية]، والهياكل العظمية للبذيرات الجيرية الصغيرة المكونة من الكالسيت، وهي نوع من الطحالب التي ازدهرت في البحار الطباشيرية.
تعتبر رواسب الطباشير في شمال غرب أوروبا من العصر الطباشيري العلوي الخاصة لصخور المجموعة الطباشيرية [الإنجليزية]، التي تشكل جروف دوفر البيضاء الواقعة على الساحل الجنوبي لإنجلترا والمنحدرات المماثلة على الساحل النورماندي الفرنسي. وتم العثور على المجموعة في إنجلترا، وشمال فرنسا والبلدان المنخفضة وشمال ألمانيا، والدنمارك وفي جوف الأجزاء الجنوبية من بحر الشمال. من صفات الطباشير أنه لا يتصلب بسهولة ولا تزال مجموعة الطباشير تتكون من رواسب فضفاضة في أماكن كثيرة. وللمجموعة أيضا مكونات أخرى مثل الحجر الجيري والأرينيت. ومن بين الأحافير التي تحتوي عليها قنافذ البحر، والسهميات والأمونيتات وزواحف بحرية مثل الموزاصور.
في جنوب أوروبا، عادة ما يكون العصر الطباشيري نظاما بحريا يتكون من طبقات جيرية صامدة أو طبقة ضعيفة من الطين الجيري. وبما أن سلاسل جبال الألب لم تتشكل بعد في العصر الطباشيري فإن هذه الرواسب تشكلت على الحافة الجنوبية للجرف القاري الأوروبي، على هامش محيط تيثس.
تسبب ركود التيارات في أعماق البحر خلال منتصف العصر الطباشيري إلى نقص الأكسجين في مياه البحر مما ترك المواد العضوية المترسبة بدون تحلل. وخلال نقص الأكسجين تركز نصف احتياطي النفط العالمي في الخليج العربي وخليج المكسيك. وخلال هذه المدة وفي أماكن كثيرة حول العالم تشكل طفل صفحي داكن عديم الأكسجين.[14] ويعتبر هذا الطفل الصفحي مصدرا مهما للنفط والغاز، كما في أعماق بحر الشمال على سبيل المثال.
الجغرافيا
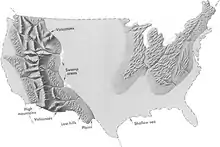
خلال العصر الطباشيري، أكملت القارة العظمى بانجيا تفككها التكتوني لتصبح قريبة من القارات الحالية، مع أختلاف كبير في مواقعها في ذلك الوقت. ومع اتساع المحيط الأطلسي، استمر التكون الجبلي الذي بدأ خلال الجوراسي في الكوردييرا الأمريكية الشمالية، كما أعقب تشكل نيفادا تشكل سيفير ولاراميد.
برغم من أن قارة غندوانا كانت سليمة في بداية العصر الطباشيري، إلا إنها انفصلت مكونة أمريكا الجنوبية، وأنتاركتيكا وابتعدت أستراليا عن أفريقيا (رغم أن الهند ومدغشقر ظلت مرتبطة ببعضهما البعض)؛ ومن هذا التباعد تشكل حديثا المحيطين الأطلسي الجنوبي والهندي. وسبب هذا التصدع النشط في ارتفاع السلاسل الجبلية الكبيرة التي تحت البحر على طول الأنقاض، رافعا مستويات البحار المتوازنة في جميع أنحاء العالم. استمر بحر تيثس في شمال أفريقيا إلى التضيق. تقدمت البحار الضحلة العريضة إلى وسط أمريكا الشمالية (الطريق البحري الغربي الداخلي) وأوروبا، ثم انحسر في وقت متأخر من هذه العصر، تاركاً وراءه رواسب بحرية كثيفة محصورة بين طبقات الفحم. في ذروة اجتياح البحر الطباشيري، غمر ما يعادل ثلث مساحة الأرض الحالية.[15]
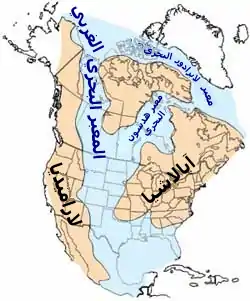
اشتهر العصر الطباشيري بصخور الطبشور؛ وفي الواقع، فإن تشكل الطباشير خلال العصر الطباشيري يعتبر أكثر من أي عصر آخر في دهر البشائر.[16] زاد نشاط نتوءات منتصف المحيط (أو بالأحرى، دوران مياه البحر عبر النتوءات المتضخمة) في دعم المحيطات بالكالسيوم. وهذا جعل المحيطات أكثر تشبعا، بالإضافة إلى زيادة التوافر البيولوجي لعنصر البذيرات الجيرية النانوية الكلسية.[17] تجعل هذه الكربونات المنتشرة وغيرها من الرواسب من سجل الصخور الطباشيري أن يكون دقيقا. تشمل التكوينات الشهيرة لأمريكا الشمالية الأحافير البحرية الغنية لتشكل طباشير سموك هيل في كانساس والحيوانات البرية في تكوين هيل كريك أواخر العصر الطباشيري . وحدثت انكشافات طباشيرية هامة أخرى في أوروبا (مثل ويلاد) وفي الصين (تكوين ييكسيان). وفي المنطقة التي أصبحت الآن الهند، انفجرت طبقات حمم ضخمة تسمى مصاطب ديكان وذلك في أواخر العصر الطباشيري وأوائل العصر الباليوسيني.
الطقس
استمر اتجاه البرودة في الفترة الأخيرة من العصر الجوراسي حتى المرحلة الأول من العصر الطباشيري. وهناك أدلة على أن تساقط الثلوج كانت كثيفة في المناطق التي فوق خطوط العرض، وأصبحت المناطق الاستوائية أكثر رطوبة مما كانت عليه خلال عصري الثلاثي والجوراسي.[18] واقتصر التجلد على الجبال الواقعة أعلى خطوط العرض، رغم أن الثلوج الموسمية قد تهطل بعيدا عن القطبين. حدث تجمع للحجارة عبر الجليد إلى البيئات البحرية خلال زمن طويل من العصر الطباشيري، لكن الأدلة على الترسب المباشر من الأنهار الجليدية مقتصر على فترة الطباشيري المبكر لحوض ايرومانغا الوقع جنوب أستراليا.[19][20]
زادت درجات الحرارة مرة أخرى بعد نهاية الحين الأول من العصر الطباشيري، وكانت هذه الظروف ثابتة حتى نهاية العصر.[18] وقد يكون سبب هذه الحرارة هو النشاط البركاني الكثيف الذي أنتج كميات كبيرة من ثاني أكسيد الكربون. تشير النسب النظائرية في الفتر ما بين بين 70-69 مليون سنة مضت و 66-65 مليون سنة مضت إلى ارتفاع ضغط ثاني أكسيد الكربون في الغلاف الجوي بمستويات 1000-1400 جزء في المليون ومتوسط درجات الحرارة السنوية في غرب تكساس بين 21 و 23 درجة مئوية (70-73 درجة فهرنهايت). وتشير علاقات ثاني أكسيد الكربون في الغلاف الجوي ودرجة الحرارة إلى أن مضاعفة ضغط ثاني أكسيد الكربون الجزئي مصحوبا بزيادة تقريبا 0.6 درجة مئوية في درجة الحرارة.[21] ويُنسب إنتاج الكميات الكبيرة من الصهارة، بشكل مختلف إلى أعمدة الوشاح أو إلى التكتونيات الممتدة،[22] التي زادت في رفع مستويات البحر إلى أعلى، بحيث غطت البحار الضحلة المناطق الواسعة من القشرة القارية. كما ساعد بحر تيثس الذي يربط المحيطات الاستوائية من الشرق إلى الغرب في تدفئة المناخ العالمي. وقد عرفت أحافير النباتات ذات التكيف الدافئ من المناطق البعيدة من شمال ألاسكا وغرينلاند، بينما تم العثور على أحافير للديناصورات في نطاق 15 درجة من القطب الجنوبي للطباشيري.[23] وهناك دليل على التجلد البحري في القطب الجنوبي في فترة التوروني.[24]
يبين التدرج اللطيف لدرجة الحرارة من خط الاستواء إلى القطبين بأن الرياح العالمية تكون أضعف، وتقود تيارات المحيطات، التي تؤدي إلى انخفاض التيار الصاعدة وركود للمحيطات أكثر مما هي عليه اليوم. ويتجلى ذلك في ترسب الطفل الصفحي المنتشر وأحداث نقص الأكسجين المتكررة.[14] تبين المادة في لب الرواسب أن درجات حرارة سطح البحر الاستوائي كانت لفترة وجيزة دافئة بقدر 42 درجة مئوية، 17 درجة مئوية، أي أكثر دفئا مما هي عليه اليوم، وأن متوسطها يبلغ حوالي 37 درجة مئوية. وفي نفس الوقت كانت درجات حرارة أعماق المحيطات تصل مابين 15 إلى 20 درجة مئوية، أي أكثر دفئا مما هي عليه اليوم.[25][26]
الحياة
النباتات

انتشرت النباتات المزهرة (كاسيات البذور) خلال هذه العصر،[27] بالرغم من أنها لم تصبح سائدة حتى مرحلة الكامباني قرب نهاية العصر. وقد ساعد ظهور النحل على تطورها؛ وفي الحقيقة، إن تطور كاسيات البذور والحشرات يعتبر مثال جيد على التطور المشترك. في نفس الوقت، استمرت بعض عاريات البذور التي كانت تعيش في حقبة الحياة الوسطى بالازدهار، ومن الأمثلة على ذلك نباتات الأروكاريا وغيرها من المخروطيات الوفيرة والمنتشرة بشكل ملحوظ. وظهرت رتب لنبات السرخس بشكل مبكر في السجل الأحفوري مثل سراخس غليكن [الإنجليزية]، وقد حققت انتشارا واسعا بشكل مبكر.[28]
حدثت بعض التغييرات على المشهد العام للطبيعة وبيئة الأرض بعد وصول كاسيات البذور. ومع ذلك فقد استمرت عاريات البذور في الهيمنة خلال العصر الطباشيري السفلي. وكانت المجموعة الوحيدة التي استمرت في الازدهار هي مجموعة الصنوبريات. وكانت السيكاديات والجنكة في تراجع فعلي، وواجهت رتب البينيتيات وجنس الهيرميرلا [الإنجليزية] انقراضا وشيكا.[29] ظهرت كاسيات البذور خلال العصر الطباشيري السفلي، وشرعت في الانتشار والتنوع التدريجي الذي قادها إلى أنواعها الحالية التي يبلغ عددها حوالي 25,0000 نوع، وعلى النقيض تبلغ الأنواع الصنوبرية الموجودة حوالي 550 نوع. وعلى عكس عاريات البذور التي تحمل البذور العارية، فإن كاسيات البذور تحصر البذور داخل المبيض، لكي تحميها من الالتهابات الفطرية، والجفاف وهجوم الحشرات. يرجع تاريخ أقدم أحافير لكاسيات البذور إلى العصر الطباشيري السفلي، أي منذ حوالي 130-120 مليون سنة مضت:وتتكون من حبوب لقاح تسمى (Clavatipollenites)، في منطقة ولدين، جنوب شرق إنجلترا. أصبحت حبوب اللقاح وأوراق الشجر مألوفة من قبل حوالي 120-100 مليون سنة. عثر على العديد من الزهور والفواكه من كاسيات البذور البدائية المحفوظة، في شرق أمريكا الشماليةٍ، وروسيا وإسرائيل، ويبدو أنها مرتبطة بنباتات المغنولية الحديثة والجميز.
ظهر خلال الطباشيري العلوي (الكامباني) تقريبا 50 فصيلة حديثة (من أصل 500)، من بينها كانت أشجار الزان، والتين، والبتولا، والبهشية، والمغنولية، والسنديان، والنخيل، والجمّيز، والجوز والصفصاف. وحبوب اللقاح والأوراق هي أكثر الأحافير شيوعًا. وقد ارتفعت مستويات حبوب لقاح كاسيات البذور من 1% من إجمالي حبوب اللقاح إلى 40% خلال فقط 20 مليون سنة، وهذا يدل على النمو الجامح للنباتات المزهرة. في البداية كانت أوراق الأشجار صغيرة وعروقها غير منتظمة ذات حواف ناعمة وبسيطة. وفي الطباشيري العلوي أصبحت أكبر وبحواف مدببة أو مفصصة، وكان عروقها ذات أنماطا أكثر انتظاما. ويعتقد أن الموائل القديمة لكاسيات البذور كانت تعاني من اضطرابات شديدة بسبب التيارات أو الفيضانات. وقد كانت الشجيرات الكبيرة والصغيرة والأعشاب السريعة النمو قادرة على السيطرة على هذه البيئات، وكانت قادرة على إنتاج البذور بسرعة. وكانت المخروطيات تعاني من مشكلة كبيرة، وهي أنه بعد التلقيح، يجب عليها الانتظار حتى يمر الشتاء قبل أن تطلق بذورها، وبالتالي فهي تحتاج عامين للإنبات. ربما يكون مرور الديناصورات المهاجرة، مفيدا لصالح الأعشاب والشجيرات لما يترتب عليه من دوس وتسميد للتربة.
تفاعل النبات والديناصور
طورت النباتات الجوراسية أنظمة دفاعية لها ضد الديناصورات النباتية المتجولة (الصربوديات والستيغوصورات) التي التي تلتهم الأجزاء العلوية من النباتات. وقد طورت عاريات البذور أشواكا أو سموما أو نكهات كريهة لحماية أنفسها من إفساد أجزاءها العليا من الرعي الجائر، وبالرغم من سلالتها على مستوى الأرض، فلم تكن بحاجة إلى مثل هذه الحماية. أدى انقراض الأنواع التي تقتات من الأجزاء العلوية من النباتات وظهور أنواع تقتات على الأجزاء السفلية منها إلى تطور النباتات القادرة على النمو والتكاثر بسرعة لإدامة أنواعها. وكانت كاسيات البذور تمتلك هذه القدرة التي لا تمتلكها عاريات البذور؛ وبالتالي تسببت ديناصورات الرعي الجديدة في ظهور كاسيات البذور. ويرى بعض الخبراء عكس ذلك، وذلك بالقول، أن انتشار كاسيات البذور كان بسبب ظهور حيوانات رعي منخفضة على حساب الرعي العالي. ولم يتم توضيح أي من النظريتين الأقرب إلى الحقيقة بعد.
التلقيح

لم تظهر النباتات المزهرة حتى حلول العصر الطباشيري، ولم تكن لديها بتلات في البداية. وقد تطورت بسبب ارتباطها بالحشرات. ويمكن القول أن الحشرات والنباتات تطورتا بشكل مشترك. ومن المحتمل أن الخنافس قامت بتلقيح بعض الخضروات التي تعتبر قريبة من أسلاف كاسيات البذور، مثل البينيتيات، التي لها أعضاء تناسلية كالزهرة التي تجذب الملقحات من خلال الرائحة العطرة أو الألوان الجذابة، وبالتالي "تدريب" بعض الحشرات للاستجابة لهذا النوع من الإغواء. قد تنتمي الحشرات الملقحة الأخرى من العصر الجوراسي والطباشيري المبكر إلى ذوات الجناحين، أو غشائيات الأجنحة أو الفراشات الليلية الصغيرة. ومن الحشرات المعروفة في العصر الطباشيري السفلي التي وجدت في مقاطعة ويلدان الإنجليزية: اليعسوب، والصراصير، والجداجد، والبق، والخنافس، والذباب، والدبابير والنمل الأبيض وغيرها. تحمل بعض غشائيات الأجنحة حبوب اللقاح داخلها، وهذا يبين أن هذا كان جزءا من نظامها الغذائي، وقد بين ظهور الشعيرات في أرجل الدبابير خلال العصر الطباشيري السفلي النشاط في التلقيح. كما أنه تم العثور على حشرات حفظت بشكل استثنائي خلال العصر الطباشيري السفلي في بايسا [الإنجليزية] (سيبيريا).
تم اكتشاف أول نحلة من العصر الطباشيري عام 2006 من قبل جورج بوانار الأبن [الإنجليزية]، وقد وجدها في كهرمان في بورما. وكانت نحلة دبور العسل البورمية [الإنجليزية]. وقد كانت تحمل مزيج من صفات النحل والدبابير.
يعتبر عنبر نيوجيرسي أحد أهم الرواسب التي تنتمي للعصر الطباشيري السفلي من مقاطعة ألافا وقد قدم منذ عام 1996 أكثر من 1500 عينة أحفورية من المفصليات، وبشكل رئيسي الحشرات (55% ذوات الجناحين، 24% غشائيات الأجنحة تشمل نحلة سريتوتريغون بريسيا [الإنجليزية] "Cretotrigona prisca"). وقد تم بالتأكيد تلقيح كاسيات البذور الأولى بواسطة حشرات مختلفة. وربما تكون العلاقات الأكثر انتقائية قد تطورت في العصر الطباشيري العلوي مع ظهور الدبابير، وكان تلقيح الزهور الصغيرة من التناظر الإشعاعي. في نهاية العصر الطباشيري وأثناء العصر الثالث، كانت الأزهار تتكيف أكثر فأكثر مع نوع واحد من الحشرات الملقحة، النحل. كان ظهور النمل الأبيض خلال العصر الطباشيري السفلي وكان ظهور النحل والنمل خلال العصر الطباشيري العلوي، مما يشير إلى التطورات الحاسمة في السلوك الاجتماعي للحشرات. وأصول هذه الحشرات الاجتماعية، ناجح جدا هذا اليوم، ويمكن أن تكون مرتبطة بالإشعاع التطوري لكاسيات البذور.
الحياة البحرية

تطورت العوالق المحيطية في نهاية العصر الطباشيري إلى خصائص حديثة تمامًا. وشهدت الدياتومات التي نشأت في العصر الجوراسي وحتى قبل ذلك بكثير بالتوسع الكبير خلال العصر الطباشيري الأوسط، جنبا إلى جنب مع السوطيات الدوارة. وحدث نفس الشيء في البحار الدافئة مع العوالق النانوية الجيرية والمنخربات عوالق (الجلوبجرينا)، التي ظهرت في العصر الجوراسي. ومن تلك اللحظة ساهمت الكميات الهائلة من العوالق النانوية في تكوين الرواسب الكلسية المعروفة باسم الطباشير. وسبح في القاع كل من الأمونيتات، والسهميات والأسماك بأنواعها المختلفة مع أجيال جديدة من الزواحف المائية. في أواخر العصر الطباشيري اختفى الإكتيوصور عمليا،[30] وأخذت مكانها أسماك القرش الكبيرة الجديدة والعظميات الضخمة مثل الزيفاكتينوس [الإنجليزية]، التي يتراوح طولها بين 2 و 4 أمتار. وهذه بدورها تتعايش مع السلاحف البحرية الكبيرة مثل الأركلون، التي تمتلك زعانف يتراوح طولها أكثر من 3 أمتار، بحيث يتجاوز طول الحيوان إذا امتدا كلاهما. وكانت الشفنينات شائعة أيضا. ومن بين الزواحف، العظايا المفلطحة، التي تنتمي إلى مجموعة البليزوصورات الطويلة العنق، التي يصل طوله إلى 12 مترا. يصل طول الموزاصور إلى 17 مترا، وكانت تعتبر أشرس الحيوانات البحرية المفترسة في العصر الطباشيري العلوي. كانت متعلقة بالسحالي الكبيرة (أعظم ما وجد على الإطلاق) المرتبطة بالثعابين الحالية (على الرغم من أنها مرتبطة بالورليات منذ فترة طويلة). وكان للموزاصور فكوك طويلة بأسنان حادة، بجانب جسمها الطويل والنحيل مع أطراف على شكل مجاديف. وكانت تتغذى على الأسماك، بالرغم من العثور على أحافير الأمونيتات عليها علامة لصف أسنان تتناسب تماما مع نمط الأسنان لبعض أنواع الموزاصورات.







 Dastilbe (شعاعيات الزعانف)
Dastilbe (شعاعيات الزعانف) Baculites (أمونيت)
Baculites (أمونيت)

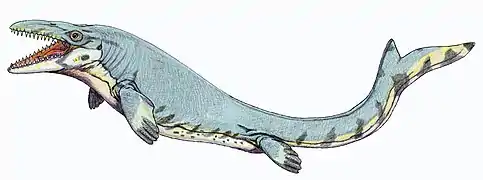

 Leptocleidus (بليزوصورات)
Leptocleidus (بليزوصورات)
 هسبرونس (الأركوصورات).
هسبرونس (الأركوصورات).
بدأت القاعيات تقدم نظرة حديثة، واستمرت عضديات الأرجل بالتراجع. وواصلت نجوم البحر والمرجانيات السداسية بالتنوع وكان للمنخربات القاعية تنوع مشابه للتنوع الحالي. وهناك توسع كبير داخل الحيوانات الحزازية في رواسب الكيلوستومات [الإنجليزية]، بأكثر من 100 نوع. انتشرت بطنيات القدم الجديدة بشكل سريع، واكتسبت جميع أعضائها تقريبا قدرات افتراسية جديدة (اختراق الأصداف وامتصاص اللحم الموجود بها، وتسميم ضحاياها، أو أبتلاع الأسماك الصغيرة). ومن الحيوانات المفترسة الأخرى القشريات، التي ظهرت في الجوراسي وهي مخلوقات تشبه الكركند الشائك وفي العصر الطباشيري تتكون من السرطان الحقيقي وجراد البحر (فتحت أصدافها بسبب كلاليبها، مثل الموجودة حالي). تقوم ذوات الصدفتين بدفن أنفسها بشكل عميق في القاع هربا من الحيوانات المفترسة، وبدأت بتطوير أصدافها لتصبح قوية أو يكون لها أشواك لردعها. كان لدى مجموعة من ذوات الصدفتين، تدعى الصدفيات القوية (Inoceramidae)، أصداف بطول 1.8 متر تقريبا. لبعض الوقت كانت كمية المرجان المطلوبة لبناء الشعاب المرجانية تحدد عمليا من قبل محار الرودست (محار ذو صدفتين مخروطيتين يصل طولها إلى متر واحد). وتثبت أنفسها على سطح صلب في القاع، وتتراكم فيما بينها وتشكل شعابا هائلة في جميع بحار العالم. يحميها هذا التشكل الكثيف من جميع الاحتمالات لالتهامها.
الحيوانات الهوائية
تطورت تيروصورات العصر الطباشيري لمجموعة متنوعة من الأشكال اتبعت اتجاهات معينة مثل:
- ازدياد الحجم خلال معظم العصر الطباشيري.
- الفقدان التدريجي للأسنان.
- تجوف العظام التي لا تخضع لجهد كبير.
هذه الصفات للتيرانودون بشكل خاص. وكانت الطيور الأحفورية للعصر الطباشيري نادرة، باستثناء الاكتشافات المعزولة من الطباشيري السفلي للطائر الصيني المكتشف في عام 1992، والأسباني (1988-1992) الإيبرومسورنيس [الإنجليزية]،[31] والكونكورنس [الإنجليزية] من الطباشيري السفلي في قونكة الإسبانية.
يعتبر المرسب الأحفوري في لياونينغ (تكوين ييكسيان) في الصين كنز لبقايا محفوظة من أنواع عديدة للديناصورات الصغيرة، والطيور والثدييات، والذي يقدم لمحة عن الحياة في العصر الطباشيري المبكر. تمثل ديناصورات السيلوروصوريات الموجودة هناك أنواع لمجموعة المانيرابتورا، والتي تشمل الطيور الحديثة وأقاربها من غير الطيور، مثل الدرومايوصوريات، والعظائيات السارقة للبيض، والتريزينوصورات، والترودونتيدات بجانب طيريات الأجنحة الأخرى. لوحظ في أحافير هذه الديناصورات التي من مرسب لياونينغ الأحفوري بوجود ريش يشبه الشعر.







الحيوانات الأرضية
كانت الثدييات صغيرة الحجم وتعتبر عنصر مهما من الحيوانات، مع تفوق أعداد عديدات الدرنات على الديناصورات في بعض الأماكن.[32] لم تكن الجرابيات الحقيقية ولا المشيميات موجودة حتى نهاية العصر،[33] لكن مجموعة متنوعة من الوحشيات البعدية الغير جرابية والوحشيات الحقيقية غير المشيميين بدأت بالتنوع بشكل كبير، من بين آكلات للحوم (Deltatheroida)، وعلّافات مائية (Stagodontidae) وعواشب (Schowalteria ،Zhelestidae). كانت المجموعات "القديمة" المختلفة مثل ثلاثية الأسنان المخروطية [الإنجليزية] شائعة في الطباشيري المبكر، ولكن كانت الحيوانات الثديية الشمالية في العصر الطباشيري المتأخر تهيمن عليها عديدات الدرنات والوحشيات، وكانت الدريولستات [الإنجليزية] تهيمن على أمريكا الجنوبية.
بالتأكيد كانت زواحف الأركوصورات، وعلى وجه الخصوص الديناصورات تحتل مكانا مهما خلال العصر الطباشيري. وفي الحقيقة كان هذا العصر الذي تم فيه تحقيق أكبر ازدهار وتطور. كانت التيروصورات شائعة في العصر الطباشيري المبكر والأوسط، لكن مع تقدم العصر بدأت بالتناقص لأسباب غير مفهومة (كان يُعتقد في السابق أنه بسبب التنافس مع الطيور البدائية، لكن الآن أصبح من المفهوم أن التشعب التكيفي للطيور لا يتوافق مع تناقص التيروصورات[34])، وبحلول نهاية العصر بقيت فصيلتان فقط.
تم العثور على مواقع الديناصورات الرئيسية في مقاطعة والدين الإنجليزية (الطباشيري السفلي) وتشكيل نامغت في منغوليا (الطباشيري العلوي)، وفي حوض نامغت في صحراء جوبي. تشمل منطقة "والدين" مساحات واسعة من الرواسب القارية والبحيرية التي تمتد في جميع أنحاء جنوب شرق إنجلترا وجزيرة وايت، رغم أنها موجودة على نطاق واسع في شمال إسبانيا. كانت الديناصورات السائدة في المنطقة من نوع طيريات القدم، وهي مجموعة ظهرت في أواخر العصر الجوراسي. وهناك أيضا عظائيات القدم، بالرغم من ندرة. تتوفر في شبه الجزيرة الإيبيرية رواسب الإيشنيت [الإنجليزية]، مع وجود بقايا غير مهمة للديناصورات، رغم ظهور العديد من الرفات مؤخرا في كل من (طرويل، وبلنسية، ولاريوخا). وقد ظهر مؤخرا في لاريوخا أحد أكبر الرواسب في العالم وآثار لكل من وحشيات الأرجل الآكلة للحوم والطيور الآكلة للأعشاب.
تنوعت الحشرات خلال العصر الطباشيري، وأقدم أنواع النمل المعروفة النمل الأبيض وظهرت بعض حرشفيات الأجنحة المعروفة، كالفراشات والعث. ظهرت حشرات المن والجنادب والزنابير.[35]
الديناصورات
وصلت الديناصورات خلال هذا الوقت إلى تشعب تكيفي كبير. وهذا هو السبب في وجود أعداد كبيرة من الأنواع ذات أنماط الحياة والأشكال المختلفة.
- طيريات الورك: حدث انتشار كبير لطيريات الورك خلال معظم العصر الطباشيري، بالإضافة إلى تنوعها من بين مختلف الحيوانات الواطنة، وهذه مؤشرات على النجاح الذي حققته. في العصر الطباشيري السفلي وجدت الإغوانادون والهيبسيلوفودون.
- الإغوانادون: وتعد أكثر انتشارا من بين طيريات القدم الكبيرة. كانت ثنائية الحركة أي على قدمين (بالرغم من وجود بعض الجدل التاريخي في هذا الصدد) وطولها 10 أمتار، تملك جمجمة تشبه جمجمة الحصان، ولها فكوك سفلية طويلة وعيون في أقصى الخلف. فكوكها مسلحة بأسنان قابلة للاستبدال، مما يبقي أسنانها حديثة. المنطقة الأمامية من الفم خالية من الأسنان ولها بنية عظمية، وأداة دقيقة لقطع الأوراق والنباتات. ويكمن التقدم الحقيقي في قدرته على المضغ. كانت تمشي على قدمين، مما يسمح لها القدرة على الوصول إلى لمناطق العليا من الأشجار، ولكن يمكنها أيضًا أن تتحرك على أربعة أرجل، انطلاقا من مورفولوجيا أيديها (مخالبها في الواقع حوافر صغيرة). مخالبها في أبهامها طويلة وحادة،[36] وكانت مختلفة جدا عن الآخرين، عندما تم العثور عليه وجد على أنفها قرن، قد يكون سلاح مفيد ضد الحيوانات المفترسة.
- الهيبسيلوفودون: يعد ثاني أكثر الديناصورات شيوعًا في مقاطعة (والدن). وقد كان حيوانا صغيرا (3 إلى 5 أمتار) له أرجل خاصة للجري بسرعة وذيل صلب يساعدها على التوازن أثناء المطاردة.
- الهادروصوريات: ديناصورات كبيرة يتراوح طولها من 10 إلى 15 أمتار، وجميعها تمتلك أجسام تعادل الشكل النموذجي لديناصورات الإغوانادونات. تتميز بالخطم المسطّح، الذي يشبه شكل منقار البط، وأسنانها مرتبة في صفوف متعددة، تصل بالغالب إلى 500 سنًا للفك السفلي. وللعديد من الهادروصورات بروز لعظم الأنف، وعَظْمُ مُقَدَّمِة الفَكِّ العُلْوِيِّ متجه للخلف، نتج عنها مجموعة من النتوءات (بعضها لوحي، والبعض الآخر أنبوبي ، والبعض الآخر بارز). يمر الهواء في الهادروصورات من قنوات أنفية وعبر حافة مقدمة الفك العلوي، التي تتعرض بالغالب من التقلبات، ووظيفة النتوءات هي توفير الإشارات المرئية والصوتية. ومن الهادروصوريات الماياصورا والباراصورولوفوس.
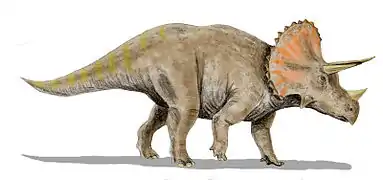
- قرنيات الوجه (السيراتوبسيدات): عاشت في أمريكا الشمالية وآسيا، ولديها رأس كبير بمنقار، وشكله من الأعلى شكل المثلث، ولديها أعدادا مختلفة من القرون والدروع العظمية التي تحمي منطقة الكتف ويشكل أيضا مغرز عضلات الفك السفلي القوية. يبلغ طول أكبر سيراتوبسيد 9 أمتار تقريا، بقرون يبلغ طولها حوالي متر واحد، ووزنه يصل إلى 6 أطنان، ومن الأمثلة عليها التريسيراتوبس، الذي وجد في غرب أمريكا الشمالية والبروتوسيراتوبس.
- الباكيسفالوصورات: وهي مجموعة أخرى من "طيريات القدم" الثنائية الأقدام ذات الجماجم السميكة. وكانت منتشرة بشكل واسع خلال العصر الطباشيري العلوي في أمريكا الشمالية وآسيا. كان الستيغوسيراس أحد أعضاء هذه الفصيلة بطول مترين فقط، بالرغم من ضخامة قبو جمجمته (25 سم، ما يقرب من نصف طول الجمجمة). يعتقد أن الباكيسفالوصورات كانت تقاتل بعضها البعض من أجل التزاوج بطريقة مماثلة لقتال الكباش وغيرها من العواشب المماثلة. ومن أبرز أجناسها الباكيسيفالوصور.
- الأنكيلوصوريات: ظهرت في العصر الجوراسي وتنوعت في العصر الطباشيري المتأخر. كانت مرتبطة ارتباطا وثيقا بالستيغوصوريات، وغالبا ما كانت كبيرة الحجم، حيث يصل طولها إلى 10 أمتار ووزنها 6 أطنان. تتميز بالدروع القوية، مع وجود صفائح عظمية على الظهر، والرقبة، والذيل والأطراف، وتعزز العظام الجمجمة، وحتى فوق العينين. كما أن لديها قرون عظمية صغيرة على الرأس وأشواك عظمية على الظهر بأنماط مختلفة. كان لدى الأنكيلوصورات المتأخرة أيضًا كتلة من العظام المدمجة في نهاية الذيل والتي تعمل بمثابة صَوْلَجان ثقيل متأرجح، بإمكانه أن يضرب أرجل الحيوانات المهاجمة بينما يبقى الحيوان ثابتا على الأرض.

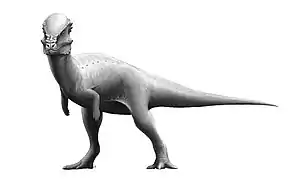

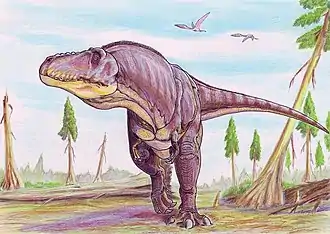
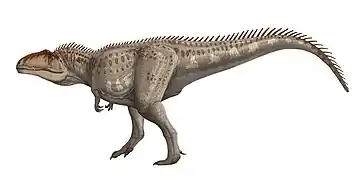
- سحليات الورك: من بين الثيروبودات الآكلة للحوم بالإمكان أن تشمل أشكال مختلفة. أدى اكتشاف الباريونيكس في عام 1983 إلى التعرف على مجموعة جديدة من الحيوانات الآكلة للحوم في هذا الحيوان، المعروف بهيكل عظمي واحد. والميزة الأبرز هي مخلب المنجلي الضخم. حيث يبلغ مخلب باريونيكس حولي 30 سم، أكثر بكثير باعتباره بطول 9 أمتار، ولم يتضح بعد ما إذا كان المخلب مثبتا باليد أو القدم. تم العثور على حراشف أسماك في معدته، مما يثبت أنه كان من آكلات الأسماك. تتميز مخالبه أنها منحنية، تساعد في التقاط الفريسة، ويبدو أنها خاصة لعدد مستقل من الثيروبودات. تشبه ديناصورات التيرانوصور ريكس (أمريكا الشمالية) والتربوصور المنغولي إلى حد كبير لسابقتها، وقد تكون من المفترسات العليا،[37] وكان التيرانوصور من أكبر الديناصورات البرية المفترسة في عصرها، بحيث تجاوزت بشكل طفيف كل من الغيغانوتوصور والسبينوصور. بلغ طول التيرانوصور 13 مترا، بارتفاع 4.8 أمتار ووزنه 5 أطنان. ولديه أسنان بطول 15 سم، أقدامه قوية وضخمة، بثلاثة أصابع في كل قدم. الذراعين تبدوان نحيلتين وقصيرة جدًا لدرجة أنها لا تصل إلى الفم، ولها أصبعين فقط. ولوحظ أن رأسه الصلب لا يقدم نظام حفظ الوزن كما في آكلات اللحوم الأخرى، ويبدو أنه مصمم للضرب، ولا يمكن لمثل هذا المفترس الكبير أن يجري بسرعة لفترة طويلة. وهو يتغذى على العواشب الكبيرة، والحيوانات ألأضعف أو التي ماتت سلفا.
- ومن آكلات اللحوم الكبيرة الأخرى الكارنوتوروس والسبينوصور. كما أنه توجد ثيروبودات مفترسة أصغر (الفيلوسيرابتور، والدينونيكوس، والأوفيرابتور، والغاليميموس، والأفيميوس [الإنجليزية]). وعلى ما يبدو أن جميع الديناصورات من فرع المانيرابتورا وربما من بعض المجموعات الأخرى من الثيروبودات كان لها ريش. وكانت الطيور هي الديناصورات الوحيدة التي نجت في نهاية العصر الطباشيري.






حدث انقراض نهاية الطباشيري
 مقالة مفصلة: انقراض العصر الطباشيري-الباليوجيني
مقالة مفصلة: انقراض العصر الطباشيري-الباليوجيني

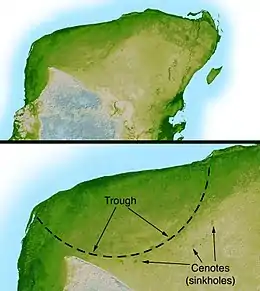
قد يكون اصطدام جسم كبير بالأرض علامة لنهاية التدهور التدريجي في التنوع البيولوجي خلال مرحلة الماسترخي من العصر الطباشيري. وكانت النتيجة هي انقراض ثلاثة أرباع أنواع النباتات والحيوانات في الأرض. أدى اصطدام إلى حدوث فاصل حاد يعرف باسم حدود الطباشيري-الباليوجيني "ط-ب" (K–Pg) (المعروفة سابقا باسم حدود K-T طباشيري-ثلاثي). وقد تطلب التنوع البيولوجي للأرض وقتا كبيرا للتعافي من هذا الحدث، بالرغم من وجود وفرة محتملة من مكامن بيئية شاغرة.[38]

كان هناك تباين كبير في معدل الانقراض بين وداخل الفروع الحيوية المختلفة بالرغم من شدة حدث انقراض الطباشيري-الباليوجيني. انخفضت أو انقرضت الأنواع التي تعتمد على التركيب الضوئي بسبب حجب جسيمات الغلاف الجوي الطاقة الشمسية. وكما هو الحال في هذا اليوم، فقد كانت كائنات التركيب الضوئي، كالعوالق النباتية والنباتات البرية تشكل جزء أساسي من السلسلة الغذائية في أواخر العصر الطباشيري، وقد عانى كل شيء يعتمد عليها. ماتت الحيوانات العاشبة التي تعتمد على النباتات والعوالق كغذاء رئيسي لها، بسبب ندرة مصادرها؛ وبناء على ذلك نفقت أكبر الحيوانات المفترسة مثل تيرانوصورات ريكس.[39]
انقرض أو تناقص كل من البذيرات الجيرية والرخويات، بما في ذلك الأمونيتات، والروديست، وحلزونات المياه العذبة وبلح البحر، بالإضافة إلى الكائنات التي تضم سلسلتها الغذائية مثل بناة الأصداف. مثلا، كان يعتقد أن الأمونيتات كانت الطعام الرئيسي لمجموعة من الزواحف البحرية العملاقة تدعى الموزاصورات والتي انقرضت عند تلك الحدود.[40]
نجى من هذه الكارثة كل من آكلات النبات والحيوانات، والحاشرات وآكلات الجيف، ويعتقد سبب ذلك هو زيادة توافر مصادر طعامها. ويبدو أنه في نهاية العصر الطباشيري لم يكن هناك ثدييات عاشبة أو آكلات لحوم. أما الثدييات والطيور التي نجت من الانقراض كانت تتغذى على الحشرات، واليرقات، والديدان والقواقع التي بدورها كانت تتغذى على البقايا النباتية والحيوانية الميتة. ويعتقد العلماء أن هذه الكائنات الحية أنقذت السلاسل الغذائية النباتية من الانهيار لأنها كانت تتغذى على الحتات.[41][38][42]
انقرضت مجموعات قليلة من حيوانات مجتمعات التيارات المائية. لأن مجتمعات التيارات المائية تعتمد بشكل أقل على طعام النباتات الحية وأكثر على الحتات التي تنغسل من الأرض، وهذا المكان الإيكولوجي بالذات منعها من الانقراض.[43] وبشكل مماثل ولكن بنمط أكثر تعقيدا في المحيطات، كان الانقراض أكثر حدة بين الحيوانات التي تعيش في العمود المائي، مقارنة بتلك الحيوانات التي تعيش في البحر أو في قاع البحر. بحيث أن الحيوانات في العمود المائي تعتمد بشكل كامل تقريبا على الإنتاج الأولي من العوالق النباتية الحية، بينما الحيوانات التي تعيش في المحيط أو في قاع المحيط تتغذى على الحتات أو يمكن أنها قد تحولت إلى كائنات تتغذى على الحتات.[38]
أكثر الناجين من الكائنات التي تتنفس الهواء، هي التماسيح والشامبسوصورات، التي كانت شبه مائية وتستطيع الوصول إلى الحتات. بإمكان التماسيح الحديثة أن تعيش كقمامات وبإمكانها أيضا أن تعيش لأشهر بدون طعام وتلجأ للسبات عندما تكون الظروف غير مواتية، وصغارها تنمو ببطء، وتتغذى في السنوات القليلة الأولى بشكل كبير على اللافقاريات والكائنات الميتة أو بقايا الكائنات الحية. وقد ارتبطت هذه الخصائص ببقاء التمساح خلال نهاية العصر الطباشيري.[41]
المراجع
ملاحظات
- ظهر مصطلح "Cretaceous" لأول مرة باللغة الإنجليزية في: "الدليل الجيولوجي" لهنري توماس دي لا بيش، (فيلادلفيا ، بنسلفانيا: كاري ولي ، 1832).من الصفحة 35: "المجموعة الرابعة. (Cretaceous) يحتوي الطباشيري على الصخور في إنجلترا وشمال فرنسا التي تتميز بالطباشير في الجزء العلوي والرمال والأحجار الرملية في الجزء السفلي."
الاقتباسات
- https://timescalefoundation.org/gssp/index.php?parentid=all
- Image:Sauerstoffgehalt-1000mj.svg
- File:OxygenLevel-1000ma.svg
- Image:Phanerozoic Carbon Dioxide.png
- Image:All palaeotemps.png
- "International chronostratigraphic chart v2018/08" (PDF) (باللغة الإنجليزية). الوسيط
|CitationClass=تم تجاهله (مساعدة). - Global Boundary Stratotype Section and Point (GSSP) of the International Commission of Stratigraphy.نسخة محفوظة 10 ديسمبر 2018 على موقع واي باك مشين.
- Glossary of Geology (الطبعة 3rd). Washington, D.C.: American Geological Institute. 1972. صفحة 165. الوسيط
|CitationClass=تم تجاهله (مساعدة) - d’Halloy, d’O., J.-J. (1822). "Observations sur un essai de carte géologique de la France, des Pays-Bas, et des contrées voisines" [Observations on a trial geological map of France, the Low Countries, and neighboring countries]. Annales des Mines. 7: 353–376. مؤرشف من الأصل في 01 أبريل 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) From page 373: "La troisième, qui correspond à ce qu'on a déja appelé formation de la craie, sera désigné par le nom de terrain crétacé." (The third, which corresponds to what was already called the "chalk formation", will be designated by the name "chalky terrain".) - Sovetskaya Enciklopediya [Great Soviet Encyclopedia] (باللغة الروسية) (الطبعة 3rd). Moscow: Sovetskaya Enciklopediya. 1974. vol. 16, p. 50. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Renne, Paul R.; et al. (2013). "Time scales of critical events around the Cretaceous-Paleogene boundary". Science. 339 (6120): 684–688. Bibcode:2013Sci...339..684R. doi:10.1126/science.1230492. PMID 23393261. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Vennari, Verónica V.; Lescano, Marina; Naipauer, Maximiliano; Aguirre-Urreta, Beatriz; Concheyro, Andrea; Schaltegger, Urs; Armstrong, Richard; Pimentel, Marcio; Ramos, Victor A. (2014). "New constraints on the Jurassic–Cretaceous boundary in the High Andes using high-precision U–Pb data". Gondwana Research. 26 (1): 374–385. Bibcode:2014GondR..26..374V. doi:10.1016/j.gr.2013.07.005. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Jaramillo, Jessica. "Entrevista al Dr. Víctor Alberto Ramos, Premio México Ciencia y Tecnología 2013" (باللغة الإسبانية). مؤرشف من الأصل في 02 أبريل 2020.
Si logramos publicar esos nuevos resultados, sería el primer paso para cambiar formalmente la edad del Jurásico-Cretácico. A partir de ahí, la Unión Internacional de la Ciencias Geológicas y la Comisión Internacional de Estratigrafía certificaría o no, depende de los resultados, ese cambio.
الوسيط|CitationClass=تم تجاهله (مساعدة) - Stanley 1999، صفحات 481–482.
- Dixon, Dougal; Benton, M J; Kingsley, Ayala; Baker, Julian (2001). Atlas of Life on Earth. New York: Barnes & Noble Books. صفحة 215. ISBN 9780760719572. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Stanley 1999، صفحة 280.
- Stanley 1999، صفحات 279–281.
- Kazlev, M.Alan. "Palaeos Mesozoic: Cretaceous: The Berriasian Age". Palaeos.com. مؤرشف من الأصل في 20 ديسمبر 2010. اطلع عليه بتاريخ 18 أكتوبر 2017. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Alley, N. F.; Frakes, L. A. (2003). "First known Cretaceous glaciation: Livingston Tillite Member of the Cadna‐owie Formation, South Australia". Australian Journal of Earth Sciences. 50 (2): 139. Bibcode:2003AuJES..50..139A. doi:10.1046/j.1440-0952.2003.00984.x. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Frakes, L. A.; Francis, J. E. (1988). "A guide to Phanerozoic cold polar climates from high-latitude ice-rafting in the Cretaceous". Nature. 333 (6173): 547. Bibcode:1988Natur.333..547F. doi:10.1038/333547a0. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Nordt, Lee; Atchley, Stacy; Dworkin, Steve (December 2003). "Terrestrial Evidence for Two Greenhouse Events in the Latest Cretaceous". GSA Today. Vol. 13 no. 12. doi:10.1130/1052-5173(2003)013<4:TEFTGE>2.0.CO;2. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Foulger, G.R. (2010). Plates vs. Plumes: A Geological Controversy. Wiley-Blackwell. ISBN 978-1-4051-6148-0. مؤرشف من الأصل في 06 أبريل 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Stanley 1999، صفحة 480–482.
- Bornemann, Norris RD; Friedrich, O; Beckmann, B; Schouten, S; Damsté, JS; Vogel, J; Hofmann, P; Wagner, T (Jan 2008). "Isotopic evidence for glaciation during the Cretaceous supergreenhouse". Science. 319 (5860): 189–92. Bibcode:2008Sci...319..189B. doi:10.1126/science.1148777. PMID 18187651. مؤرشف من الأصل في 06 أبريل 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Warmer than a Hot Tub: Atlantic Ocean Temperatures Much Higher in the Past" فيز. Retrieved 12/3/06. نسخة محفوظة 2020-04-08 على موقع واي باك مشين.
- Skinner & Porter 1995، صفحة 557.
- Coiro, Mario; Doyle, James A.; Hilton, Jason (2019-01-25). "How deep is the conflict between molecular and fossil evidence on the age of angiosperms?". New Phytologist (باللغة الإنجليزية). 223 (1): 83–99. doi:10.1111/nph.15708. PMID 30681148. الوسيط
|CitationClass=تم تجاهله (مساعدة) - C.Michael Hogan. 2010. Fern. Encyclopedia of Earth. National council for Science and the Environment نسخة محفوظة November 9, 2011, على موقع واي باك مشين.. Washington, DC
- "Introduction to the Bennettitales". University of California Museum of Paleontology. مؤرشف من الأصل في 08 أبريل 2020. اطلع عليه بتاريخ 30 مايو 2014. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Paleobiology / Smithsonian National Museum of Natural History" en. مؤرشف من الأصل في 12 أبريل 2020. اطلع عليه بتاريخ 13 أبريل 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة); Invalid|script-title=: missing prefix (مساعدة) - Museo de las Ciencias de Castilla-La Mancha. "Museo de las Ciencias". مؤرشف من الأصل في 11 de noviembre de 2007. اطلع عليه بتاريخ 31 de diciembre de 2007. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ الوصول=, |تاريخ أرشيف=(مساعدة) - Kielan-Jaworowska, Zofia; Cifelli, Richard L.; Luo, Zhe-Xi (2005). Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. Columbia University Press. صفحة 299. ISBN 9780231119184. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Halliday, Thomas John Dixon; Upchurch, Paul; Goswami, Anjali (29 June 2016). "Eutherians experienced elevated evolutionary rates in the immediate aftermath of the Cretaceous–Palaeogene mass extinction". Proc. R. Soc. B. 283 (1833): 20153026. doi:10.1098/rspb.2015.3026. PMC 4936024. PMID 27358361. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wilton, Mark P. (2013). Pterosaurs: Natural History, Evolution, Anatomy. Princeton University Press. ISBN 978-0691150611. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Life of the Cretaceous". www.ucmp.Berkeley.edu. مؤرشف من الأصل في 14 أبريل 2020. اطلع عليه بتاريخ 18 أكتوبر 2017. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Senckenberg, forschungsinstitut und naturmuseum. "Iguanodon". مؤرشف من الأصل في 13 de febrero de 2009. اطلع عليه بتاريخ 31 de diciembre de 2007. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ الوصول=, |تاريخ أرشيف=(مساعدة) - Universidad de Berkeley. "The Tyrannosauridae". مؤرشف من الأصل في 26 يوليو 2019. اطلع عليه بتاريخ 31 de diciembre de 2007. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ الوصول=(مساعدة) - MacLeod, N; Rawson, PF; Forey, PL; Banner, FT; Boudagher-Fadel, MK; Bown, PR; Burnett, JA; et al. (1997). "The Cretaceous–Tertiary biotic transition". Journal of the Geological Society. 154 (2): 265–292. Bibcode:1997JGSoc.154..265M. doi:10.1144/gsjgs.154.2.0265. مؤرشف من الأصل في 18 أبريل 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wilf, P; Johnson KR (2004). "Land plant extinction at the end of the Cretaceous: a quantitative analysis of the North Dakota megafloral record". Paleobiology. 30 (3): 347–368. doi:10.1666/0094-8373(2004)030<0347:LPEATE>2.0.CO;2. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Kauffman, E (2004). "Mosasaur Predation on Upper Cretaceous Nautiloids and Ammonites from the United States Pacific Coast". PALAIOS. 19 (1): 96–100. doi:10.1669/0883-1351(2004)019<0096:MPOUCN>2.0.CO;2. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Shehan, P; Hansen, TA (1986). "Detritus feeding as a buffer to extinction at the end of the Cretaceous". Geology. 14 (10): 868–870. Bibcode:1986Geo....14..868S. doi:10.1130/0091-7613(1986)14<868:DFAABT>2.0.CO;2. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Aberhan, M; Weidemeyer, S; Kieesling, W; Scasso, RA & Medina, FA (2007). "Faunal evidence for reduced productivity and uncoordinated recovery in Southern Hemisphere Cretaceous–Paleogene boundary sections". Geology. 35 (3): 227–230. Bibcode:2007Geo....35..227A. doi:10.1130/G23197A.1. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Sheehan, PM; Fastovsky, DE (1992). "Major extinctions of land-dwelling vertebrates at the Cretaceous–Paleogene boundary, eastern Montana". Geology. 20 (6): 556–560. Bibcode:1992Geo....20..556S. doi:10.1130/0091-7613(1992)020<0556:MEOLDV>2.3.CO;2. الوسيط
|CitationClass=تم تجاهله (مساعدة)
قائمة المراجع
- Yuichiro Kashiyama; Nanako O. Ogawa; Junichiro Kuroda; Motoo Shiro; Shinya Nomoto; Ryuji Tada; Hiroshi Kitazato; Naohiko Ohkouchi (May 2008). "Diazotrophic cyanobacteria as the major photoautotrophs during mid-Cretaceous oceanic anoxic events: Nitrogen and carbon isotopic evidence from sedimentary porphyrin". Organic Geochemistry. 39 (5): 532–549. doi:10.1016/j.orggeochem.2007.11.010. الوسيط
|CitationClass=تم تجاهله (مساعدة)CS1 maint: ref=harv (link) - Larson, Neal L; Jorgensen, Steven D; Farrar, Robert A; Larson, Peter L (1997). Ammonites and the other Cephalopods of the Pierre Seaway. Geoscience Press. الوسيط
|CitationClass=تم تجاهله (مساعدة)CS1 maint: ref=harv (link) - Ogg, Jim (June 2004). "Overview of Global Boundary Stratotype Sections and Points (GSSP's)". مؤرشف من الأصل في 16 يوليو 2006. الوسيط
|CitationClass=تم تجاهله (مساعدة)CS1 maint: ref=harv (link) - Ovechkina, M.N.; Alekseev, A.S. (2005). "Quantitative changes of calcareous nannoflora in the Saratov region (Russian Platform) during the late Maastrichtian warming event" (PDF). Journal of Iberian Geology. 31 (1): 149–165. مؤرشف من الأصل (PDF) في 24 أغسطس 2006. الوسيط
|CitationClass=تم تجاهله (مساعدة)CS1 maint: ref=harv (link) - Rasnitsyn, A.P.; Quicke, D.L.J. (2002). History of Insects. سبرنجر. ISBN 978-1-4020-0026-3. الوسيط
|CitationClass=تم تجاهله (مساعدة)CS1 maint: ref=harv (link)—detailed coverage of various aspects of the evolutionary history of the insects. - Skinner, Brian J.; Porter, Stephen C. (1995). The Dynamic Earth: An Introduction to Physical Geology (الطبعة 3rd). New York: John Wiley & Sons. ISBN 0-471-60618-9. الوسيط
|CitationClass=تم تجاهله (مساعدة)CS1 maint: ref=harv (link) - Stanley, Steven M. (1999). Earth System History. New York: W.H. Freeman and Company. ISBN 0-7167-2882-6. الوسيط
|CitationClass=تم تجاهله (مساعدة)CS1 maint: ref=harv (link) - Taylor, P. D.; Wilson, M. A. (2003). "Palaeoecology and evolution of marine hard substrate communities". Earth-Science Reviews. 62 (1): 1–103. Bibcode:2003ESRv...62....1T. doi:10.1016/S0012-8252(02)00131-9. الوسيط
|CitationClass=تم تجاهله (مساعدة)
| العصر الطباشيري | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| المبكر | المتأخر | ||||||||||
| البرياسي | الفالانجيني | الهاتريفي | الباريمي | الأبتي | الألبي | السينوماني | التوروني | الكونياكي | السانتوني | الكامباني | الماسترخي |
| دهر البشائر | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| حقبة الحياة القديمة | حقبة الحياة الوسطى | حقبة الحياة الحديثة | |||||||||
| الكامبري | الأوردفيشي | السيلوري | الديفوني | الفحمي | البرمي | الثلاثي | الجوراسي | الطباشيري | الباليوجين | النيوجيني | الرباعي |
 بوابة علم طبقات الأرض
بوابة علم طبقات الأرض بوابة علوم الأرض
بوابة علوم الأرض بوابة ديناصورات
بوابة ديناصورات بوابة علم الأحياء القديمة
بوابة علم الأحياء القديمة بوابة زمن
بوابة زمن