مضاد السموم (نظام)
نظام مضاد السموم (بالإنجليزية: Toxin-antitoxin system) هو عبارة عن مجموعة من اثنين أو أكثر من الجينات المرتبطة ارتباطًا وثيقًا والتي تعمل معًا على ترميز كل من بروتين "السموم" و "مضاد السموم" المقابل. يتم توزيع أنظمة مضادات السموم على نطاق واسع في بدائيات النوى ، وغالبًا ما تمتلكها الكائنات الحية في نسخ متعددة. عندما يتم احتواء هذه الأنظمة على البلازميدات - عناصر وراثية قابلة للتحويل - فإنها تضمن بقاء الخلايا الوليدة التي ترث البلازميد فقط بعد انقسام الخلية. إذا كان البلازميد غائبًا في الخلية الوليدة ، يتحلل الترياق غير المستقر ويقتل البروتين السام المستقر الخلية الجديدة ؛ يُعرف هذا باسم "القتل بعد الفصل العنصري". [2]
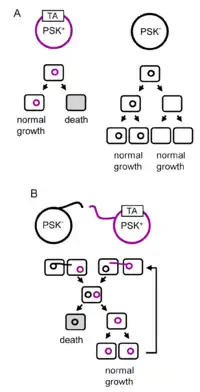
عادةً ما يتم تصنيف أنظمة مضادات السموم وفقًا لكيفية تحييد مضاد السموم من السم. في نظام مضاد للسموم من النوع الأول ، يتم منع ترجمة الحمض النووي الريبي المرسال (mRNA) الذي يشفر السم عن طريق ارتباط مضاد سموم الحمض النووي الريبي الصغير غير المشفر الذي يربط توكسين الرنا المرسال. يتم تثبيط البروتين السام في نظام النوع الثاني بعد التحويل عن طريق الارتباط ببروتين مضاد السموم. تتكون أنظمة مضادات السموم من النوع الثالث من رنا صغير يرتبط مباشرة ببروتين السم ويثبط نشاطه. هناك أيضًا أنواع من الرابع إلى السادس ، وهي أقل شيوعًا. غالبًا ما يتم توريث جينات مضادات السموم من خلال النقل الأفقي للجينات وترتبط بالبكتيريا المسببة للأمراض ، حيث تم العثور عليها في البلازميدات التي تمنح مقاومة المضادات الحيوية والفوعة.[3]
توجد أيضًا أنظمة مضادات السموم الصبغية ، والتي يُعتقد أن بعضها يؤدي وظائف خلوية مثل الاستجابة للضغوط ، والتسبب في توقف دورة الخلية والتسبب في موت الخلية المبرمج. من الناحية التطورية ، يمكن اعتبار أنظمة مضادات السموم DNA أنانية من حيث أن الغرض من الأنظمة هو التكاثر ، بغض النظر عما إذا كانت تفيد الكائن الحي المضيف أم لا. اقترح البعض نظريات تكيفية لشرح تطور أنظمة التكسينات المضادة للسموم. على سبيل المثال ، من الممكن أن تكون أنظمة مضادات السموم الصبغية قد تطورت لمنع وراثة عمليات حذف كبيرة من جينوم المضيف. تحتوي أنظمة مضادات السموم على العديد من تطبيقات التكنولوجيا الحيوية ، مثل الحفاظ على البلازميدات في خطوط الخلايا ، وأهداف المضادات الحيوية ، ونواقل الاختيار الإيجابية.[4]
وظائفه البيولوجية
وتتميز أنظمة السموم المضادة للوسم جيدا بوحدات الإدمان للبلازميدات . واقتُرح أيضاً أن تكون نظم التكسينات المضادة للسموم قد تطورت كنماذج استبعادية من البلازميد. خلية من شأنها أن تحمل اثنين من البلازميدات من نفس المجموعة عدم التوافق في نهاية المطاف توليد اثنين من بنات الخلايا التي تحمل إما plasmid. وينبغي أن واحدة من هذه البلازميدات ترميز لنظام TA، "التشريد" من قبل نظام آخر TA خالية من البلازميد سيمنع ميراثها وبالتالي الحث على القتل بعد الفصل العنصري. وقد تم تأكيد هذه النظرية من خلال نمذجة الكمبيوتر. ويمكن أيضاً العثور على نظم مضادة للسموم على عناصر وراثية متنقلة أخرى مثل المواثرات المترافقة والبكترية المعتدلة ويمكن أن تكون متورطة في صيانة هذه العناصر ومنافسة تلك العناصر.
يمكن أن تمنع أنظمة مضادات السموم الحذف الكبير الضار في الجينوم البكتيري ، على الرغم من أن الحذف من مناطق الترميز الكبيرة قاتلة لخلية الابنة بغض النظر عن ذلك. وفي فيبريو الكوليرا، تبين أن نظماً متعددة من النوع الثاني للسموم المضادة للسموم الموجودة في إنتيغورون فائقة المفعول تمنع فقدان الكاسيتات الجينية.
استقرار ولياقة الحمض النووي المتحرك
كما هو مذكور أعلاه ، تتميز أنظمة مضادات السموم بأنها وحدات إدمان البلازميد. تم اقتراح أن أنظمة مضادات السموم قد تطورت كوحدات استبعاد البلازميد. الخلية التي ستحمل بلازميدين من نفس مجموعة عدم التوافق ستنتج في النهاية خليتين من بنات تحمل أيًا من البلازميد. إذا تم ترميز أحد هذه البلازميدات لنظام TA ، فإن "إزاحته" بواسطة نظام بلازميد آخر خالٍ من TA سيمنع توريثه وبالتالي يؤدي إلى القتل بعد الفصل العنصري. [5] تم تأكيد هذه النظرية من خلال النمذجة الحاسوبية . [6] ويمكن أيضا أنظمة السامة الترياق يمكن العثور على غيرها من العناصر الجينية المحمولة مثل اقترانية ترانسبوزونات والمعتدلة البكتيريا ويمكن أن تكون متورطة في صيانة والمنافسة من هذه العناصر. [7]
استقرار الجينوم
يمكن أن تمنع أنظمة مضادات السموم عمليات الحذف الكبيرة الضارة في الجينوم البكتيري ، على الرغم من أنه يمكن القول إن حذف مناطق الترميز الكبيرة يكون قاتلًا للخلية الوليدة بغض النظر. [10] في Vibrio cholerae ، تم إثبات أن العديد من أنظمة مضادات السموم من النوع الثاني الموجودة في super-Integron تمنع فقدان أشرطة الجينات. [11]
موت الخلية الإيثاري
mazEF ، وهو موضع مضاد للسموم موجود في E. coli والبكتيريا الأخرى للحث على موت الخلايا المبرمج استجابة للجوع ، وتحديداً نقص الأحماض الأمينية . [12] سيؤدي هذا إلى إطلاق محتويات الخلية لامتصاصها من قبل الخلايا المجاورة ، مما يحتمل أن يمنع موت الأقارب ، وبالتالي زيادة اللياقة الشاملة للخلية التي هلكت. سيكون هذا مثالًا على الإيثار وكيف يمكن للمستعمرات البكتيرية أن تشبه الكائنات متعددة الخلايا . [6] ومع ذلك ، فإن "PCD بوساطة mazEF " تم دحضه إلى حد كبير من خلال العديد من الدراسات. [13] [14] [15]
تحمل الاجهاد
تنص نظرية أخرى على أن أنظمة مضادات السموم الصبغية مصممة لتكون جراثيم بدلاً من جراثيم . [16] RelE ، على سبيل المثال ، هو مثبط عالمي للترجمة ، يتم إحداثه أثناء إجهاد المغذيات . من خلال إيقاف الترجمة تحت الضغط ، يمكن أن تقلل من فرصة المجاعة عن طريق خفض متطلبات المغذيات للخلية. [17] ومع ذلك ، فقد تبين أن العديد من أنظمة مضادات السموم ، بما في ذلك relBE ، لا تعطي أي ميزة تنافسية تحت أي ظروف إجهاد. [14]
مكافحة الإدمان
لقد تم اقتراح أن متماثلات الكروموسومات لأنظمة مضادات السموم البلازميدية قد تعمل كوحدات مضادة للإدمان ، والتي من شأنها أن تسمح للنسل بفقدان البلازميد دون التعرض لتأثيرات السم الذي يشفره. [18] على سبيل المثال ، يمكن لنسخة كروموسومية من مضاد سم ccdA المشفر في كروموسوم Erwinia chrysanthemi تحييد سم ccdB المشفر على البلازميد F وبالتالي منع تنشيط السموم عند فقد هذا البلازميد. [19] وبالمثل ، فإن مضاد السموم ataR المشفر على كروموسوم الإشريكية القولونية O157: H7 قادر على تحييد ذيفان ataT P المشفر على البلازميدات الموجودة في الإشريكية القولونية المعوية الأخرى. [20]
حماية فايج
تم إثبات أن أنظمة مضادات السموم من النوع الثالث (AbiQ) تحمي البكتيريا من العاثيات بإيثار. [21] [22] أثناء العدوى ، تقوم العاثيات باختطاف النسخ والترجمة ، مما قد يمنع تجديد مضادات السموم ويطلق السم ، مما يؤدي إلى ما يسمى "عدوى فاشلة". [21] [22] وقد لوحظت تأثيرات وقائية مماثلة مع النوع الأول ، [23] النوع الثاني ، [24] والنوع الرابع (أبي إي) [25] أنظمة التوكسين المضادة للسموم.
يمكن أن يحدث البدء الفاشل (Abi) أيضًا بدون أنظمة مضادات السموم ، ويوجد العديد من بروتينات Abi من الأنواع الأخرى. تعمل هذه الآلية على وقف تكرار العاثيات ، وحماية السكان بشكل عام من الأذى. [26]
ثبات مضادات الميكروبات
عندما يتم تحدي البكتيريا بالمضادات الحيوية ، فإن مجموعة سكانية فرعية صغيرة ومتميزة من الخلايا قادرة على تحمل العلاج من خلال ظاهرة يطلق عليها اسم "المثابرة" (لا ينبغي الخلط بينها وبين المقاومة ). [27] نظرًا لخصائصها المضادة للجراثيم ، فقد كان يُعتقد سابقًا أن أنظمة مضادات السموم من النوع الثاني مسؤولة عن الثبات ، عن طريق تحويل جزء صغير من السكان البكتيرية إلى حالة نائمة. [28] ومع ذلك ، فقد تم إبطال هذه الفرضية على نطاق واسع. [29] [30] [31]
الحمض النووي الأناني
تم استخدام أنظمة مضادات السموم كأمثلة للحمض النووي الأناني كجزء من وجهة نظر التطور الجينية . لقد تم الافتراض بأن مواقع مضادات السموم تعمل فقط للحفاظ على الحمض النووي الخاص بها ، على حساب الكائن الحي المضيف. [1] [32] وبالتالي ، فإن أنظمة مضادات السموم الصبغية لن تؤدي أي غرض ويمكن معاملتها على أنها "دنا غير مرغوب فيه". على سبيل المثال ، تبين أن نظام ccdAB المشفر في كروموسوم الإشريكية القولونية O157: H7 يخضع للاختيار السلبي ، وإن كان بمعدل بطيء بسبب خصائصه المسببة للإدمان. [33]
أنواع النظام
النوع الأول
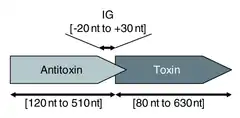
تعتمد أنظمة مضادات السموم من النوع الأول على الاقتران الأساسي بين الحمض النووي الريبي المضاد التكميلي مع مرنا التوكسين. يتم بعد ذلك منع ترجمة mRNA إما عن طريق التحلل عبر RNase III أو بإغلاق تسلسل Shine-Dalgarno أو موقع ربط الريبوسوم الخاص بالسموم mRNA. غالبًا ما يتم ترميز السم ومضاد السموم على خيوط متقابلة من الحمض النووي. المنطقة المتداخلة 5 ' أو 3' بين الجينين هي المنطقة المشاركة في الاقتران الأساسي التكميلي ، عادةً ما بين 19-23 زوجًا من القواعد المتجاورة. [34]
السموم من النوع الأول عبارة عن بروتينات صغيرة كارهة للماء تمنح السمية عن طريق إتلاف أغشية الخلايا . [1] تم تحديد عدد قليل من الأهداف داخل الخلايا من السموم من النوع الأول ، ربما بسبب الطبيعة الصعبة لتحليل البروتينات السامة لمضيفيها البكتيريين. [35]
تتضمن أنظمة النوع الأول أحيانًا مكونًا ثالثًا. في حالة نظام hok / sok ذي الخصائص الجيدة ، بالإضافة إلى hokoxin وsok antitoxin ، هناك جين ثالث يسمى mok . يتداخل إطار القراءة المفتوح هذا بشكل كامل تقريبًا مع إطار التوكسين ، وتعتمد ترجمة السم على ترجمة هذا المكون الثالث. [36] وبالتالي ، فإن ارتباط مضاد السموم بالسم يكون أحيانًا تبسيطًا ، وفي الواقع يرتبط مضاد السموم بحمض نووي ثالث ، والذي يؤثر بعد ذلك على ترجمة السموم. [34]
أمثلة على الأنظمة
| توكسين | أنتيتوكسين | ملاحظات | المرجع. |
|---|---|---|---|
| hok | sok | النظام الأصلي والأكثر فهماً من النوع الأول المضاد للسموم (في الصورة) ، والذي يعمل على تثبيت البلازميدات في عدد من البكتيريا سالبة الجرام | [34] |
| fst | محدد الاستقرار الاسمي RNAII | أول نظام من النوع الأول يتم التعرف عليه في البكتيريا موجبة الجرام | [37] |
| نظام مضادات السموم TisB-IstR | نظام مضادات السموم TisB-IstR | نظام كروموسومي مستحث في استجابة SOS | [38] |
| ldrD | rdld | نظام الكروموسومات في Enterobacteriaceae | [39] |
| نظام مضاد السموم FlmA-FlmB | flmB | متماثل hok / sok ، والذي يعمل أيضًا على استقرار بلازميد F | [40] |
| متلازمة القولون المتهيج | sib | اكتشف هذا المضاد في المناطق الجينية للإشريكية القولونية ، وكان يسمى في الأصل QUAD RNA | [41] |
| txpA / brnT | ratA | يضمن وراثة عنصر الجلد أثناء التبويض في العصوية الرقيقة | [42] |
| symE | symR | نظام كروموسومي مستحث في استجابة SOS | [43] |
| XCV2162 | ptaRNA1 | نظام تم تحديده في Xanthomonas campestris مع توزيع غير منتظم للتطور. | [44] |
| timP | timR | تم تحديد نظام كروموسومي في السالمونيلا | [45] |
| aapA1 | ISOA1 | وحدة TA من النوع 1 في Helicobacter pylori | [46] |
| sprA1 | sprA1as | تقع داخل جزيرة S. aureus الصغيرة المُمرضة (SaPI). أكواد SprA1 لببتيد صغير سام للخلايا ، PepA1 ، الذي يعطل أغشية بكتريا S. aureus وكريات الدم الحمراء المضيفة. | [47] [48] |
النوع الثاني
اكتب II أنظمة السامة الترياق عادة ما تكون مفهومة، أفضل من نوع I. [34] في هذا النظام عطوب proteic الترياق بإحكام الربط ويحول دون نشاط السم مستقرة. [35] أكبر عائلة من أنظمة مضاد السموم من النوع الثاني هي vapBC ، [49] والتي تم العثور عليها من خلال أبحاث المعلوماتية الحيوية لتمثل ما بين 37 و 42 ٪ من جميع مواقع النوع الثاني المتوقعة. [8] [9] يتم تنظيم أنظمة النوع الثاني في عوامل مع وجود البروتين المضاد للسموم عادةً في الجزء العلوي من السم ، مما يساعد على منع التعبير عن السم بدون مضاد السم. [50] يبلغ طول البروتينات عادةً حوالي 100 حمض أميني ، [34] وتظهر سمية بعدة طرق: يؤثر CcdB ، على سبيل المثال ، على تكرار الحمض النووي عن طريق تسميم الحمض النووي gyrase [51] حين أن سموم MazF و RelE هي نوكلياز داخلية تشق mRNAs الخلوية في زخارف تسلسل محدد. [52] [17] النشاط السام الأكثر شيوعًا هو البروتين الذي يعمل بمثابة نوكلياز داخلي ، يُعرف أيضًا باسم interferase (التداخل) . [53] [54]
إحدى السمات الرئيسية لاتفاقيات المساعدة التقنية هي التنظيم الذاتي. يرتبط مركب البروتين المضاد للسموم والسموم بالمشغل الموجود في بداية جينات TA. ينتج عن هذا قمع مشغل TA. مفتاح التنظيم هو (1) الترجمة التفاضلية لبروتينات TA و (2) التحلل البروتيني التفاضلي لبروتينات TA. كما أوضح " نموذج الترجمة-reponsive " ، [55] درجة التعبير تتناسب عكسًا مع تركيز مجمع TA القمعي. يتناسب تركيز مركب TA بشكل مباشر مع معدل الترجمة العالمي. كلما ارتفع معدل الترجمة زاد تعقيد TA وأقل نسخ TA mRNA. خفض معدل الترجمة ، وأقل من مركب TA وأعلى التعبير. ومن ثم ، فإن التعبير النسخي لأوبرون TA يتناسب عكسيا مع معدل الترجمة.
يمكن أحيانًا مشاركة بروتين ثالث في أنظمة مضاد السموم من النوع الثاني. في حالة نظام ω-ε-ζ (omega-epsilon-zeta) ، فإن بروتين أوميغا هو بروتين مرتبط بالحمض النووي ينظم بشكل سلبي نسخ النظام بأكمله. [56] وبالمثل ، فإن بروتين paaR2 ينظم التعبير عن نظام التوكسين المضاد للسموم paaR2-paaA2-parE2 . [57] يمكن العثور على أنظمة مضادات السموم الأخرى مع مرافق كمكون ثالث. [58] هذا المرافِق ضروري للطي المناسب لمضاد السموم ، مما يجعل الترياق مدمنًا على مرافقه المشابه.
أمثلة على الأنظمة
| توكسين | أنتيتوكسين | ملاحظات | المرجع. |
|---|---|---|---|
| نظام مكافحة السموم من النوع الثاني CcdA / CcdB | نظام مكافحة السموم من النوع الثاني CcdA / CcdB | وجدت على البلازميد F من الإشريكية القولونية | [51] |
| نظام مضاد السموم من النوع الثاني ParDE | نظام مضاد السموم من النوع الثاني ParDE | وجدت في نسخ متعددة في Caulobacter crescentus | [59] |
| mazF | mazE | توجد في E. coli وفي كروموسومات بكتيريا أخرى | [23] |
| yafO | yafN | نظام ناتج عن استجابة SOS لتلف الحمض النووي في E. coli | [60] |
| hica | hicB | وجدت في العتائق والبكتيريا | [61] |
| kid | kis | يثبت البلازميد R1 ويرتبط بنظام CcdB / A | [16] |
| ζ | ε | توجد في الغالب في البكتيريا موجبة الجرام | [56] |
| ataT | ataR | توجد في E. coli و Klebsiella spp. | [62] |
النوع الثالث
| ToxN_toxin (مادة سامة) | |
|---|---|
| معرف | |
| رمز | ToxiN هو النوع الثالث من نظام مضاد السموم |
| قاعدة بيانات عوائل البروتينات | PF13958 |
تعتمد أنظمة مضادات السموم من النوع الثالث على التفاعل المباشر بين البروتين السام ومضاد سم الحمض النووي الريبي. يتم تحييد الآثار السامة للبروتين بواسطة جين RNA. [63] أحد الأمثلة على ذلك هو نظام ToxIN من مسببات الأمراض البكتيرية Erwinia carotovora . يبلغ طول بروتين ToxN السام حوالي 170 من الأحماض الأمينية وقد ثبت أنه سام للإشريكية القولونية . تم تثبيط النشاط السام لـ ToxN بواسطة ToxI RNA ، وهو RNA مع 5.5 تكرار مباشر لعزر نيوكليوتيد 36 (AGGTGATTTGCTACCTTTAAGTGCAGCTAGAAATTC). [21] [64] وجد التحليل البلوري لـ ToxIN أن تثبيط ToxN يتطلب تكوين مركب ToxIN ثلاثي ، حيث تربط ثلاثة مونومرات ToxI ثلاثة مونومرات ToxN ؛ يتم تجميع المركب معًا عن طريق تفاعلات بروتينية واسعة النطاق. [65]
النوع الرابع
تشبه أنظمة مضادات السموم من النوع الرابع أنظمة النوع الثاني ، لأنها تتكون من بروتينين. على عكس أنظمة النوع الثاني ، فإن مضاد السم في أنظمة مضاد السموم من النوع الرابع يقاوم نشاط السم ، ولا يتفاعل البروتينان بشكل مباشر. [66] [67]
النوع الخامس
ghoST هو نظام مضاد للسموم من النوع V ، حيث يشق مضاد السموم (GhoS) ghoT mRNA. يتم تنظيم هذا النظام بواسطة نظام النوع الثاني ، mqsRA . [68]
تطبيقات التكنولوجيا الحيوية
بدأت تطبيقات التكنولوجيا الحيوية لأنظمة التوكسين المضادة للسموم تتحقق من قبل العديد من منظمات التكنولوجيا الحيوية. [70] [16] الاستخدام الأساسي هو الحفاظ على البلازميدات في مزرعة خلايا بكتيرية كبيرة. في تجربة لفحص فعالية موقع hok / sok ، وجد أن الاستقرار الفصل للبلازميد المُدرج الذي يعبر عن بيتا غالاكتوزيداز قد زاد بنسبة تتراوح بين 8 و 22 مرة مقارنة بثقافة تحكم تفتقر إلى نظام مضاد السموم. [71] [72] في نطاق واسع الكائنات الحية الدقيقة عمليات مثل التخمير ، وخلايا ذرية تفتقر البلازميد إدراج غالبا ما يكون أعلى لياقة بدنية من أولئك الذين يرثون البلازميد ويمكن تكومبيتي الكائنات الحية الدقيقة مرغوب فيه. يحافظ نظام مضاد السموم على البلازميد وبالتالي يحافظ على كفاءة العملية الصناعية. [70]
بالإضافة إلى ذلك ، قد تكون أنظمة مضادات السموم هدفًا مستقبليًا للمضادات الحيوية . يمكن أن يساعد تحريض وحدات انتحارية ضد مسببات الأمراض في مكافحة المشكلة المتنامية لمقاومة الأدوية المتعددة . [73]
ضمان قبول البلازميد للإدخال مشكلة شائعة في استنساخ الحمض النووي. يمكن استخدام أنظمة مضادات السموم للاختيار الإيجابي فقط لتلك الخلايا التي تناولت بلازميدًا يحتوي على الجين المُدخَل محل الاهتمام ، وفحص تلك التي تفتقر إلى الجين المُدخل. مثال على هذا التطبيق يأتي من التوكسين المشفر ccdB ، والذي تم دمجه في ناقلات البلازميد . [74] ثم يتم استهداف الجين المعني لإعادة الاتحاد في موضع ccdB ، مما يؤدي إلى تعطيل نسخ البروتين السام. وبالتالي ، فإن الخلايا التي تحتوي على البلازميد ولكن لا تحتوي على الملحق ، تموت بسبب التأثيرات السامة لبروتين CcdB ، وتبقى فقط تلك التي تحتوي على الملحق. [70]
تطبيق مثال آخر يتضمن كلا من توكسين CcdB ومضاد سم CcdA. تم العثور على CcdB في الجينوم البكتيري المؤتلف ويتم إدخال نسخة معطلة من CcdA في ناقل بلازميد خطي . يتم إضافة تسلسل إضافي قصير إلى الجين المعني الذي ينشط الترياق عند حدوث الإدخال. هذا الأسلوب يضمن التوجه محددة الإدراج الجينات. [74]
يجب احتواء الكائنات المعدلة وراثيًا في منطقة محددة مسبقًا أثناء البحث . [73] يمكن أن تتسبب أنظمة مضادات السموم في انتحار الخلايا في ظروف معينة ، مثل الافتقار إلى وسيط نمو خاص بالمختبر لن يصادفوه خارج نطاق المختبر الخاضع للرقابة. [16] [75]
انظر أيضًا
مراجع
- Rosenberg, Susan M., المحرر (March 2009). "Bacterial toxin-antitoxin systems: more than selfish entities?". PLOS Genetics. 5 (3): e1000437. doi:10.1371/journal.pgen.1000437. PMID 19325885. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ramisetty BC, Raj S, Ghosh D (December 2016). "Escherichia coli MazEF toxin-antitoxin system does not mediate programmed cell death". Journal of Basic Microbiology. 56 (12): 1398–1402. doi:10.1002/jobm.201600247. PMID 27259116. ^ Jump up to: a b c d
- Diago-Navarro E, Hernandez-Arriaga AM, López-Villarejo J, Muñoz-Gómez AJ, Kamphuis MB, Boelens R, Lemonnier M, Díaz-Orejas R (August 2010). "parD toxin-antitoxin system of plasmid R1--basic contributions, biotechnological applications and relationships with closely-related toxin-antitoxin systems". The FEBS Journal. 277 (15): 3097–117. doi:10.1111/j.1742-4658.2010.07722.x. PMID 20569269. ^ Jump up to: a b
- Fineran PC, Blower TR, Foulds IJ, Humphreys DP, Lilley KS, Salmond GP (January 2009). "The phage abortive infection system, ToxIN, functions as a protein-RNA toxin-antitoxin pair". Proceedings of the National Academy of Sciences of the United States of America. 106 (3): 894–9. Bibcode:2009PNAS..106..894F. doi:10.1073/pnas.0808832106. PMC 2630095. PMID 19124776. ^ Jump up to: a b
- "Postsegregational killing does not increase plasmid stability but acts to mediate the exclusion of competing plasmids". Proceedings of the National Academy of Sciences of the United States of America. 97 (23): 12643–8. November 2000. Bibcode:2000PNAS...9712643C. doi:10.1073/pnas.220077897. PMID 11058151. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Genetic addiction: selfish gene's strategy for symbiosis in the genome". Genetics. 172 (2): 1309–23. February 2006. doi:10.1534/genetics.105.042895. PMID 16299387. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Hypothetical functions of toxin-antitoxin systems". Journal of Bacteriology. 189 (17): 6089–92. September 2007. doi:10.1128/JB.00958-07. PMID 17616596. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Toxin-antitoxin loci are highly abundant in free-living but lost from host-associated prokaryotes". Nucleic Acids Research. 33 (3): 966–76. 2005. doi:10.1093/nar/gki201. PMID 15718296. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "RASTA-Bacteria: a web-based tool for identifying toxin-antitoxin loci in prokaryotes". Genome Biology. 8 (8): R155. 2007. doi:10.1186/gb-2007-8-8-r155. PMID 17678530. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Comparative analysis of superintegrons: engineering extensive genetic diversity in the Vibrionaceae". Genome Research. 13 (3): 428–42. March 2003. doi:10.1101/gr.617103. PMID 12618374. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Chromosomal toxin-antitoxin loci can diminish large-scale genome reductions in the absence of selection". Molecular Microbiology. 63 (6): 1588–605. March 2007. doi:10.1111/j.1365-2958.2007.05613.x. PMID 17367382. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "An Escherichia coli chromosomal "addiction module" regulated by guanosine [corrected] 3',5'-bispyrophosphate: a model for programmed bacterial cell death". Proceedings of the National Academy of Sciences of the United States of America. 93 (12): 6059–63. June 1996. Bibcode:1996PNAS...93.6059A. doi:10.1073/pnas.93.12.6059. PMID 8650219. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "mazEF-mediated programmed cell death in bacteria: "what is this?"". Critical Reviews in Microbiology. 41 (1): 89–100. February 2015. doi:10.3109/1040841X.2013.804030. PMID 23799870. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "What is the benefit to Escherichia coli of having multiple toxin-antitoxin systems in its genome?". Journal of Bacteriology. 189 (17): 6101–8. September 2007. doi:10.1128/JB.00527-07. PMID 17513477. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Escherichia coli MazEF toxin-antitoxin system does not mediate programmed cell death". Journal of Basic Microbiology. 56 (12): 1398–1402. December 2016. doi:10.1002/jobm.201600247. PMID 27259116. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "parD toxin-antitoxin system of plasmid R1--basic contributions, biotechnological applications and relationships with closely-related toxin-antitoxin systems". The FEBS Journal. 277 (15): 3097–117. August 2010. doi:10.1111/j.1742-4658.2010.07722.x. PMID 20569269. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "RelE, a global inhibitor of translation, is activated during nutritional stress". Proceedings of the National Academy of Sciences of the United States of America. 98 (25): 14328–33. December 2001. Bibcode:2001PNAS...9814328C. doi:10.1073/pnas.251327898. PMID 11717402. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Horizontal gene transfer of chromosomal Type II toxin-antitoxin systems of Escherichia coli". FEMS Microbiology Letters. 363 (3): fnv238. February 2016. doi:10.1093/femsle/fnv238. PMID 26667220. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Chromosomal toxin-antitoxin systems may act as antiaddiction modules". Journal of Bacteriology. 190 (13): 4603–9. July 2008. doi:10.1128/JB.00357-08. PMID 18441063. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Novel toxins from type II toxin-antitoxin systems with acetyltransferase activity". Plasmid. 93: 30–35. September 2017. doi:10.1016/j.plasmid.2017.08.005. PMID 28941941. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The phage abortive infection system, ToxIN, functions as a protein-RNA toxin-antitoxin pair". Proceedings of the National Academy of Sciences of the United States of America. 106 (3): 894–9. January 2009. Bibcode:2009PNAS..106..894F. doi:10.1073/pnas.0808832106. PMID 19124776. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "AbiQ, an abortive infection mechanism from Lactococcus lactis". Applied and Environmental Microbiology. 64 (12): 4748–56. December 1998. doi:10.1128/AEM.64.12.4748-4756.1998. PMID 9835558. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Escherichia coli mazEF-mediated cell death as a defense mechanism that inhibits the spread of phage P1". Molecular Genetics and Genomics. 272 (2): 227–34. September 2004. doi:10.1007/s00438-004-1048-y. PMID 15316771. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Exclusion of T4 phage by the hok/sok killer locus from plasmid R1". Journal of Bacteriology. 178 (7): 2044–50. April 1996. doi:10.1128/jb.178.7.2044-2050.1996. PMID 8606182. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "A widespread bacteriophage abortive infection system functions through a Type IV toxin-antitoxin mechanism". Nucleic Acids Research. 42 (7): 4590–605. April 2014. doi:10.1093/nar/gkt1419. PMID 24465005. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Seed, KD (June 2015). "Battling Phages: How Bacteria Defend against Viral Attack". PLOS Pathogens. 11 (6): e1004847. doi:10.1371/journal.ppat.1004847. PMID 26066799. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Bacterial persistence: a model of survival in changing environments". Genetics. 169 (4): 1807–14. April 2005. doi:10.1534/genetics.104.035352. PMID 15687275. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Molecular mechanisms underlying bacterial persisters". Cell. 157 (3): 539–48. April 2014. doi:10.1016/j.cell.2014.02.050. PMID 24766804. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "What Is the Link between Stringent Response, Endoribonuclease Encoding Type II Toxin-Antitoxin Systems and Persistence?". Frontiers in Microbiology (باللغة الإنجليزية). 7: 1882. 2016. doi:10.3389/fmicb.2016.01882. PMID 27933045. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Prophages and Growth Dynamics Confound Experimental Results with Antibiotic-Tolerant Persister Cells". mBio. 8 (6): e01964–17. December 2017. doi:10.1128/mBio.01964-17. PMID 29233898. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Reassessing the Role of Type II Toxin-Antitoxin Systems in Formation of Escherichia coli Type II Persister Cells". mBio. 9 (3): e00640–18. June 2018. doi:10.1128/mBio.00640-18. PMID 29895634. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Endoribonuclease type II toxin-antitoxin systems: functional or selfish?". Microbiology. 163 (7): 931–939. July 2017. doi:10.1099/mic.0.000487. PMID 28691660. مؤرشف من الأصل في 25 يناير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The decay of the chromosomally encoded ccdO157 toxin-antitoxin system in the Escherichia coli species". Genetics. 181 (4): 1557–66. April 2009. doi:10.1534/genetics.108.095190. PMID 19189956. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Small toxic proteins and the antisense RNAs that repress them". Microbiology and Molecular Biology Reviews. 72 (4): 579–89, Table of Contents. December 2008. doi:10.1128/MMBR.00025-08. PMID 19052321. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Toxins-antitoxins: plasmid maintenance, programmed cell death, and cell cycle arrest". Science. 301 (5639): 1496–9. September 2003. Bibcode:2003Sci...301.1496H. doi:10.1126/science.1088157. PMID 12970556. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Competitive inhibition of natural antisense Sok-RNA interactions activates Hok-mediated cell killing in Escherichia coli". Nucleic Acids Research. 34 (20): 5915–22. 2006. doi:10.1093/nar/gkl750. PMID 17065468. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The antisense RNA of the par locus of pAD1 regulates the expression of a 33-amino-acid toxic peptide by an unusual mechanism". Molecular Microbiology. 37 (3): 652–60. August 2000. doi:10.1046/j.1365-2958.2000.02035.x. PMID 10931358. الوسيط
|CitationClass=تم تجاهله (مساعدة) (الاشتراك مطلوب) - "The small RNA IstR inhibits synthesis of an SOS-induced toxic peptide". Current Biology. 14 (24): 2271–6. December 2004. doi:10.1016/j.cub.2004.12.003. PMID 15620655. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Molecular characterization of long direct repeat (LDR) sequences expressing a stable mRNA encoding for a 35-amino-acid cell-killing peptide and a cis-encoded small antisense RNA in Escherichia coli". Molecular Microbiology. 45 (2): 333–49. July 2002. doi:10.1046/j.1365-2958.2002.03042.x. PMID 12123448. الوسيط
|CitationClass=تم تجاهله (مساعدة) (الاشتراك مطلوب) - "Nucleotide sequence and transcriptional analysis of a third function (Flm) involved in F-plasmid maintenance". Gene. 66 (2): 259–68. June 1988. doi:10.1016/0378-1119(88)90362-9. PMID 3049248. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Repression of small toxic protein synthesis by the Sib and OhsC small RNAs". Molecular Microbiology. 70 (5): 1076–93. December 2008. doi:10.1111/j.1365-2958.2008.06394.x. PMID 18710431. الوسيط
|CitationClass=تم تجاهله (مساعدة) (الاشتراك مطلوب) - "Small untranslated RNA antitoxin in Bacillus subtilis". Journal of Bacteriology. 187 (19): 6641–50. October 2005. doi:10.1128/JB.187.19.6641-6650.2005. PMID 16166525. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "RNA antitoxins". Current Opinion in Microbiology. 10 (2): 117–24. April 2007. doi:10.1016/j.mib.2007.03.003. PMID 17376733. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "A novel family of plasmid-transferred anti-sense ncRNAs". RNA Biology. 7 (2): 120–4. March 2010. doi:10.4161/rna.7.2.11184. PMID 20220307. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Salmonella Protein TimP Targets the Cytoplasmic Membrane and Is Repressed by the Small RNA TimR". mBio. 11 (6): e01659–20, /mbio/11/6/mBio.01659–20.atom. November 2020. doi:10.1128/mBio.01659-20. PMID 33172998 تأكد من صحة قيمة
|pmid=(مساعدة). الوسيط|CitationClass=تم تجاهله (مساعدة) - "Mechanistic insights into type I toxin antitoxin systems in Helicobacter pylori: the importance of mRNA folding in controlling toxin expression". Nucleic Acids Research. 45 (8): 4782–4795. May 2017. doi:10.1093/nar/gkw1343. PMID 28077560. مؤرشف من الأصل في 2 يونيو 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "A cis-antisense RNA acts in trans in Staphylococcus aureus to control translation of a human cytolytic peptide". Nature Structural & Molecular Biology. 19 (1): 105–12. December 2011. doi:10.1038/nsmb.2193. PMID 22198463. مؤرشف من الأصل في 26 أبريل 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Functional and structural insights of a Staphylococcus aureus apoptotic-like membrane peptide from a toxin-antitoxin module". The Journal of Biological Chemistry. 287 (52): 43454–63. December 2012. doi:10.1074/jbc.M112.402693. PMID 23129767. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The vapBC operon from Mycobacterium smegmatis is an autoregulated toxin-antitoxin module that controls growth via inhibition of translation". Journal of Molecular Biology. 390 (3): 353–67. July 2009. doi:10.1016/j.jmb.2009.05.006. PMID 19445953. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Mechanisms for Differential Protein Production in Toxin-Antitoxin Systems". Toxins. 9 (7): 211. July 2017. doi:10.3390/toxins9070211. PMID 28677629. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Cell killing by the F plasmid CcdB protein involves poisoning of DNA-topoisomerase II complexes" (PDF). Journal of Molecular Biology. 226 (3): 735–45. August 1992. doi:10.1016/0022-2836(92)90629-X. PMID 1324324. مؤرشف من الأصل (PDF) في 27 أكتوبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "MazF cleaves cellular mRNAs specifically at ACA to block protein synthesis in Escherichia coli". Molecular Cell. 12 (4): 913–23. October 2003. doi:10.1016/S1097-2765(03)00402-7. PMID 14580342. الوسيط
|CitationClass=تم تجاهله (مساعدة) - RNA decay by messenger RNA interferases. 447. 2008. صفحات 521–35. doi:10.1016/S0076-6879(08)02225-8. ISBN 978-0-12-374377-0. PMID 19161859. الوسيط
|CitationClass=تم تجاهله (مساعدة) - mRNA interferases, sequence-specific endoribonucleases from the toxin-antitoxin systems. 85. 2009. صفحات 467–500. doi:10.1016/S0079-6603(08)00812-X. ISBN 978-0-12-374761-7. PMID 19215780. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ramisetty, Bhaskar Chandra Mohan (2020). "Regulation of Type II Toxin-Antitoxin Systems: The Translation-Responsive Model". Frontiers in Microbiology (باللغة الإنجليزية). 11. doi:10.3389/fmicb.2020.00895. ISSN 1664-302X. مؤرشف من الأصل في 23 يونيو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "ε/ζ systems: their role in resistance, virulence, and their potential for antibiotic development". Journal of Molecular Medicine. 89 (12): 1183–94. December 2011. doi:10.1007/s00109-011-0797-4. PMID 21822621. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "New toxins homologous to ParE belonging to three-component toxin-antitoxin systems in Escherichia coli O157:H7" (PDF). Molecular Microbiology. 76 (3): 719–32. May 2010. doi:10.1111/j.1365-2958.2010.07129.x. PMID 20345661. مؤرشف من الأصل (PDF) في 11 مايو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "SecB-like chaperone controls a toxin-antitoxin stress-responsive system in Mycobacterium tuberculosis". Proceedings of the National Academy of Sciences of the United States of America. 108 (20): 8438–43. May 2011. Bibcode:2011PNAS..108.8438B. doi:10.1073/pnas.1101189108. PMID 21536872. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Interaction specificity, toxicity and regulation of a paralogous set of ParE/RelE-family toxin-antitoxin systems". Molecular Microbiology. 77 (1): 236–51. July 2010. doi:10.1111/j.1365-2958.2010.07207.x. PMID 20487277. الوسيط
|CitationClass=تم تجاهله (مساعدة) (الاشتراك مطلوب) - "An SOS-regulated type 2 toxin-antitoxin system". Journal of Bacteriology. 191 (24): 7456–65. December 2009. doi:10.1128/JB.00963-09. PMID 19837801. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "HicA of Escherichia coli defines a novel family of translation-independent mRNA interferases in bacteria and archaea". Journal of Bacteriology. 191 (4): 1191–9. February 2009. doi:10.1128/JB.01013-08. PMID 19060138. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "fMet" (PDF). Nature Chemical Biology. 13 (6): 640–646. June 2017. doi:10.1038/nchembio.2346. PMID 28369041. مؤرشف من الأصل (PDF) في 30 نوفمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Bacteriophage resistance mechanisms". Nature Reviews. Microbiology. 8 (5): 317–27. May 2010. doi:10.1038/nrmicro2315. PMID 20348932. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Mutagenesis and functional characterization of the RNA and protein components of the toxIN abortive infection and toxin-antitoxin locus of Erwinia". Journal of Bacteriology. 191 (19): 6029–39. October 2009. doi:10.1128/JB.00720-09. PMID 19633081. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "A processed noncoding RNA regulates an altruistic bacterial antiviral system". Nature Structural & Molecular Biology. 18 (2): 185–90. February 2011. doi:10.1038/nsmb.1981. PMID 21240270. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "A novel family of Escherichia coli toxin-antitoxin gene pairs". Journal of Bacteriology. 185 (22): 6600–8. November 2003. doi:10.1128/jb.185.22.6600-6608.2003. PMID 14594833. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The Toxin-Antitoxin System DarTG Catalyzes Reversible ADP-Ribosylation of DNA". Molecular Cell. 64 (6): 1109–1116. December 2016. doi:10.1016/j.molcel.2016.11.014. PMID 27939941. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Type II toxin/antitoxin MqsR/MqsA controls type V toxin/antitoxin GhoT/GhoS". Environmental Microbiology. 15 (6): 1734–44. June 2013. doi:10.1111/1462-2920.12063. PMID 23289863. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "A bacterial toxin inhibits DNA replication elongation through a direct interaction with the β sliding clamp". Molecular Cell. 52 (5): 617–28. December 2013. doi:10.1016/j.molcel.2013.10.014. PMID 24239291. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The art of selective killing: plasmid toxin/antitoxin systems and their technological applications". BioTechniques. 45 (3): 344–6. September 2008. doi:10.2144/000112955. PMID 18778262. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Temperature and growth rate effects on the hok/sok killer locus for enhanced plasmid stability". Biotechnology Progress. 10 (6): 621–9. 1994. doi:10.1021/bp00030a600. PMID 7765697. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Combining the hok/sok, parDE, and pnd postsegregational killer loci to enhance plasmid stability". Applied and Environmental Microbiology. 63 (5): 1917–24. May 1997. doi:10.1128/AEM.63.5.1917-1924.1997. PMID 9143123. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Prokaryotic toxin-antitoxin stress response loci". Nature Reviews. Microbiology. 3 (5): 371–82. May 2005. doi:10.1038/nrmicro1147. PMID 15864262. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Positive-selection vectors using the F plasmid ccdB killer gene". Gene. 148 (1): 71–4. October 1994. doi:10.1016/0378-1119(94)90235-6. PMID 7926841. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "A dual lethal system to enhance containment of recombinant micro-organisms". Microbiology. 149 (Pt 12): 3595–601. December 2003. doi:10.1099/mic.0.26618-0. PMID 14663091. الوسيط
|CitationClass=تم تجاهله (مساعدة)
روابط خارجية
- راستا - المسح الآلي السريع للسموم ومضادات السموم في البكتيريا.
 بوابة طب
بوابة طب بوابة صحة
بوابة صحة.svg.png.webp) بوابة علم الأحياء الدقيقة
بوابة علم الأحياء الدقيقة بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي
| مقالة «مضاد السموم (نظام)» أُنشئت ضمن الدورة الأولى من مشروع المعرفة التي أُجريت طوال شهر ديسمبر 2020. |