إنزيمات الاقتطاع
إنزيمات الاقتطاع (بالإنجليزية: restiction enzymes أو restriction endonuclease) أو إنزيمات القطع أو الإنزيمات القاطعة أو إنزيمات التقييد هي الإنزيمات التي تقطع تتاليات ال DNA عند مواضع محددة تعرف بمقرات الاقتطاع، وتوجد إنزيمات الاقتطاع لدى الجراثيم الذي يعتقد بأنه يقيها من غزو العاثيات وتستخدم تلك الإنزيمات كأدوات في الدراسات الوراثية والتشخيص الوراثي.[1][2][3]
التاريخ
نشأ مصطلح إنزيم الاقتطاع من دراسات عاثية لامدا (Lambda phage)، وهو فيروس يصيب البكتيريا، وظاهرة اقتطاع وتعديل العاثيات التي يتحكم فيها المضيف. حصل التعرف على هذه الظاهرة لأول مرة في العمل المنجز في مختبرات سلفادور لوريا (Salvador Luria) وجين ويجل (Jean Weigle) وجوزيبي بيرتاني (Giuseppe Bertani) في أوائل الخمسينيات من القرن الماضي. وُجِد أنه، بالنسبة للعاثية لامدا التي يمكن أن تنمو بشكل جيد في سلالة واحدة من الإشريكية القولونية، على سبيل المثال الإشريكية القولونية سي، عندما تنمو في سلالة أخرى، على سبيل المثال الإشريكية القولونية ك، يمكن أن تنخفض محاصيلها بشكل كبير، بقدر 3-5 أوامر من حيث الحجم. تُعرف الخلية المضيفة، في هذا المثال الإشريكية القولونية ك، بمُضيف الاقتطاع ويبدو أن لديه القدرة على تقليل النشاط البيولوجي للعاثية لامدا. إذا أُسِّسَت العاثية في سلالة واحدة، فإن قدرة تلك العاثية على النمو تصبح أيضًا مقيدة في سلالات أخرى. في الستينيات من القرن العشرين، ظهر في العمل المُنجَز في مختبرات فرنر أربر (Werner Arber) وماثيو ميسيلسون (Matthew Meselson) أن التقييد ناتج عن الانقسام الأنزيمي للحمض النووي للعاثية، لذلك فإن الإنزيم المعنيّ كان يسمى إنزيم التقييد (الاقتطاع). [4][5][6][7][8][9]
كانت إنزيمات الاقتطاع التي درسها أربر وميسيلسون هي إنزيمات الاقتطاع من النوع الأول، والتي تشق الحمض النووي بشكل عشوائي بعيدًا عن موقع التعرف. في عام 1970، قام كلٌ من هاملتون سميث (Hamilton Othanel Smith)، وتوماس جاي كيلي (Thomas J. Kelly)، وكينت ويلكوكس (Kent Wilcox) بعزل وتمييز أول إنزيم اقتطاع من النوع الثاني (HindII) من بكتيريا المستدمية النزلية. تعد إنزيمات الاقتطاع من هذا النوع أكثر فائدة للعمل المخبري لأنها تشق الحمض النووي في موقع تسلسل التعرف الخاص بها وهي الأكثر استخدامًا كأداة بيولوجيا جزيئية. في وقت لاحق، أظهر دانيال ناثانز (Daniel Nathans) وكاثلين دانا (Kathleen Danna) أن انقسام الحمض النووي لفيروس القرد 40 عن طريق إنزيمات الاقتطاع ينتج شظايا مُحدَّدة يمكن فصلها باستخدام الرحلان الكهربائي باستخدام عديد الأكريلاميد، مما يدل على أنه يمكن أيضًا استخدام إنزيمات الاقتطاع لرسم خرائط الحمض النووي. من أجل عملهم في اكتشاف وتوصيف إنزيمات الاقتطاع، مُنحت جائزة نوبل عام 1978 في علم وظائف الأعضاء أو الطب لكل من فرنر أربر ودانيال ناثانز وهاملتون سميث. يسمح اكتشاف إنزيمات الاقتطاع بمعالجة الحمض النووي، مما يؤدي إلى تطوير تقنية الحمض النووي المؤتلف التي لها العديد من التطبيقات، على سبيل المثال، السماح بإنتاج البروتينات على نطاق واسع مثل الإنسولين البشري الذي يستخدمه مرضى السكري. [10][11][12][13][14][15][16][17]
التسمية
منذ اكتشافها في السبعينيات من القرن العشرين، حصل التعرف على العديد من إنزيمات الاقتطاع، على سبيل المثال، حصل تمييز أكثر من 3500 إنزيم اقتطاع مختلف من النوع الثاني. يُسمى كل إنزيم على اسم البكتيريا التي عُزِل منها، وذلك باستخدام نظام تسمية يعتمد على الجنس والنوع والسلالة البكتيرية. [18][19][20]
الأصول
من المُحتمل أن تكون إنزيمات الاقتطاع قد تطوّرت من سلف مشترك وانتشرت عبر نقل الجينات الأفقي.[21][22] بالإضافة إلى ذلك، هناك أدلة متزايدة على أن النوكليازات الداخلية للاقتطاع تطوّرت كعنصر جيني أناني. [23]
موضع القطع
إنزيمات القطع تتعرف على تتابع محدد من النيوكليوتيدات وتنتج قطع مزدوج في الـDNA. يكون طول التتابع في العادة ما بين 4 إلى 8 نيوكليوتيدات. كل إنزيم قاطع يعتبر عن مقص خاص لقطع الدي إن أي في نقطة محددة.و يتعرف الإنزيم القاطع على مكان القطع حسب تسلسل الدي إن أي للقطع؛ فكل إنزيم قاطع يقطع في تسلسل محدد.فمثلا الإنزيم القاطع المعروف بالهيبا وأحد(Hpa I )يقطع عندما يجد 6 من الأحماض النووية في هذا التسلسل ( GTTAAC) بينما الإنزيم القاطع إيكو أر واحد (Eco RI ) يقطع عندما يجد 6 من الأحماض النووية في هذا التسلسل( GATATC).و للمعلومية فإن هيبا واحد سمي بهذا الاسم لأنه يوجد في بكتيريا الهيموفلس بارا انفلونزا (Hemophilus parainfluenzae ) و هذا الإنزيم يعتبر من الإنزيمات التي تقطع بشكل رأسي مستقيم. بينما إنزيم الإيكو آر واحد فهو مأخوذ من بكتيريا الإيكو كولي (Escherichia coli )، ويعتبر من الإنزيمات التي تقطع بشكل متعرج.
الأنواع
تُصنَّف النوكليازات الداخلية للاقتطاع الطبيعية إلى أربع مجموعات (النوع 1 و2 و3 و4) بناءً على تكوينها ومتطلبات إنزيم العامل المساعد، وطبيعة تسلسلها المستهدف، وموقع انقسام الحمض النووي بالنسبة للتسلسل المستهدف.[24][25][26] ومع ذلك، تُظهِر تحليلات تسلسل الحمض النووي لإنزيمات الاقتطاع اختلافات كبيرة، مما يشير إلى وجود أكثر من أربعة أنواع.[27] تتعرف جميع أنواع الإنزيمات على تسلسلات حمض نووي قصيرة محددة وتُجري انقسام النواة للحمض النووي لإعطاء شظايا محددة باستخدام محطة 5-الفوسفات. وهي تختلف في تسلسل التعرف (Recognition sequence)، وتكوين الوحدة الفرعية، وموضع الانقسام، ومتطلبات العامل المساعد،[28][29] على النحو المُلخص أدناه:
- تلتصق إنزيمات النوع 1 (رقم التصنيف الإنزيمي 3.1.21.3) في مواقع بعيدة عن موقع التعرف، تتطلب كلاً من أدينوسين ثلاثي الفوسفات وS-أدينوسيل الميثيونين لتعمل، بروتين متعدد الوظائف مع كل من اهتضام التقييد وأنشطة ناقلة الميثيل (Methylase) (رقم التصنيف الإنزيمي 2.1.1.72).
- تنقسم إنزيمات النوع 2 (رقم التصنيف الإنزيمي 3.1.21.4) داخل أو على مسافات قصيرة محددة من موقع التعرف، معظمهم يتطلب المغنيسيوم، لديهم وظيفة واحدة (اهتضام التقييد) وهي إنزيمات مستقلة عن ناقلة الميثيل.
- تنقسم إنزيمات النوع 3 (رقم التصنيف الإنزيمي 3.1.21.5) في مواقع على مسافة قصيرة من موقع التعرف، تتطلب أدينوسين ثلاثي الفوسفات (لكن لا تتحلل بالماء)، يُحفِّز S-أدينوسيل الميثيونين التفاعل ولكنه غير مطلوب، إنها موجودة كجزء من مركب مع تعديل ناقلة الميثيل (رقم التصنيف الإنزيمي 2.1.1.72).
- تستهدف إنزيمات النوع 4 الحمض النووي المُعدَّل، على سبيل المثال المُمَثيَل (Methylated)، هيدروكسي مُمثيل (Hydroxymethylated) وجليكوزيل هيدروكسي مُمثيل (Glucosyl-Hydroxymethylated) الحمض النووي.
- تستخدم إنزيمات النوع 5 الحمض النووي الريبوزي الإرشادي (gRNAs).
النوع 1
كانت إنزيمات الاقتطاع من النوع الأول هي الأولى التي حُدِّدت، وحُدِّدت لأول مرة في سلالتين مختلفتين (K-12 و B) من الإشريكية القولونية. تقطع هذه الإنزيمات في موقع مختلف، وعلى بعد مسافة عشوائية (1000 زوج قاعدي (bp) على الأقل) من موقع التعرف عليها. يتبع الانقسام في هذه المواقع العشوائية عملية نقل الحمض النووي، والتي تُظهِر أن هذه الإنزيمات هي أيضًا محركات جزيئية. موقع التعرف غير متماثل ويتكون من جزأين محددين، أحدهما يحتوي على 3-4 نيوكليوتيدات والآخر يحتوي على 4-5 نيوكليوتيدات، مفصولة بفاصل غير محدد من حوالي 6-8 نيوكليوتيدات. هذه الإنزيمات متعددة الوظائف وقادرة على كل من اهتضام التقييد وأنشطة التعديل، اعتمادًا على حالة المثيلة للحمض النووي المُستهدَف. العوامل المساعدة S-أدينوسيل الميثيونين، والأدينوسين ثلاثي الفوسفات المُتحلل، وأيونات المغنيسيوم مطلوبة لنشاطهم الكامل. تمتلك إنزيمات الاقتطاع من النوع الأول ثلاث وحدات فرعية تسمى HsdR و HsdM و HsdS. الأول مطلوب لاهتضام التقييد، ويعد الثاني ضروريًا لإضافة مجموعات الميثيل إلى الحمض النووي المضيف (نشاط ناقلة الميثيل)، والثالث مهم لخصوصية موقع التعرف (ربط الحمض النووي) بالإضافة إلى كل من اهتضام التقييد (انقسام الحمض النووي) ونشاط التعديل (دنا ميتيل-ترانسفيراز). [24][30]
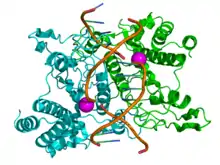
النوع 2
تختلف إنزيمات الاقتطاع النموذجية من النوع الثاني عن إنزيمات الاقتطاع من النوع الأول بطرق متعددة. إنها تشكل ثنائيات متجانسات، مع مواقع التعرف التي عادة ما تكون غير مُجزَّأة ومتجانسة وطول 4-8 نيوكليوتيدات. إنهم يتعرفون على الحمض النووي ويقطعونه في نفس الموقع، ولا يستخدمون أدينوسين ثلاثي الفوسفات أو S-أدينوسيل الميثيونين لنشاطهم، وعادة ما يحتاجون فقط المغنيسيوم كعامل مساعد. تقطع هذه الإنزيمات رابطة فوسفات ثنائي الأستر للحلزون المزدوج للحمض النووي. يمكن أن تقطع إما في مركز كلا الجدولين للحصول على نهاية حادة، أو في وضع متعرج تاركًا نتوءات تسمى النهايات اللاصقة (Sticky ends).[32] هذه هي أنزيمات الاقتطاع الأكثر شيوعًا واستخدامًا. في التسعينيات من القرن العشرين وأوائل العقد الأول من القرن الحادي والعشرين، اكتُشفَت إنزيمات جديدة من هذه العائلة لا تتبع جميع المعايير الكلاسيكية لفئة الإنزيم هذه، وطُوِّرَت تسميات فصيلة فرعية جديدة لتقسيم هذه العائلة الكبيرة إلى فئات فرعية بناءً على الانحرافات عن الخصائص النموذجية لإنزيمات النوع الثاني. تُعرَّف هذه المجموعات الفرعية باستخدام لاحقة حرف.
إنزيمات الاقتطاع من النوع 2 ب (على سبيل المثال، BcgI و BplI) متعددة المرات (Multimers)، وتحتوي على أكثر من وحدة فرعية واحدة. إنهم يقطعون الحمض النووي على كلا الجانبين من التعرف على قطع موقع التعرف. تتطلب كلًا من العوامل المساعدة لـ S-أدينوسيل الميثيونين والمغنيسيوم . نوكليازات الاقتطاع من نوع 2 هـ (على سبيل المثال، NaeI) تقطع الحمض النووي بعد التفاعل مع نسختين من تسلسل التعرف عليها. يعمل أحد مواقع التعرف كهدف للقطع، بينما يعمل الآخر كتفارغية تعمل على تسريع أو تحسين كفاءة قطع الإنزيم. على غرار إنزيمات النوع 2 هـ، تتفاعل نوكليازات تقييد النوع 2 F (على سبيل المثال NgoMIV) مع نسختين من تسلسل التعرف الخاص بها ولكنها تقطع كلا التسلسل في نفس الوقت. تحتوي نوكليازات الاقتطاع من النوع 2 جي (على سبيل المثال، RM.Eco57I) على وحدة فرعية واحدة، مثل إنزيمات اقتطاع النوع الثاني الكلاسيكية، ولكنها تتطلب أن يكون العامل المساعد S-أدينوسيل الميثيونين نشطًا. نوكليازات تقييد النوع 2 م (مثل DpnI) قادرة على التعرف على مثيلة الحمض النووي وقطعه.[33][34] نوكليازات اقتطاع نوع 2 س (على سبيل المثال، FokI) تقطع الحمض النووي على مسافة مُحددة من مواقع التعرف غير المتماثلة وغير المتناظرة، تُستخدَم هذه الخاصية على نطاق واسع لأداء تقنيات الاستنساخ في المختبر مثل استنساخ البوابة الذهبية (Golden Gate Cloning). قد تعمل هذه الإنزيمات كموانع. وبالمثل، تتكون إنزيمات اقتطاع النوع 2 تي (على سبيل المثال، Bpu10I و BslI) من وحدتين فرعيتين مختلفتين، يتعرف البعض على التسلسلات المتناظرة بينما يمتلك البعض الآخر مواقع تمييز غير متماثلة. [35]
النوع 3
تتعرف إنزيمات الاقتطاع من النوع الثالث (على سبيل المثال، EcoP15) على تسلسلين منفصلين غير متماثلين مُوجَّهين بشكل عكسي. قاموا بقطع الحمض النووي حوالي 20-30 زوجًا قاعديًا بعد موقع التعرف. تحتوي هذه الإنزيمات على أكثر من وحدة فرعية وتتطلب عوامل مساعدة S-أدينوسيل الميثيونين وأدينوسين ثلاثي الفوسفات لدورها في مثيلة الحمض النووي واهتضام التقييد، على التوالي. إنها مكونات آليات تعديل اقتطاع الحمض النووي بدائي النواة الذي يحمي الكائن الحي من غزو الحمض النووي الغريب. إنزيمات النوع الثالث عبارة عن بروتينات مُغايرة للقليل من البروتينات متعددة الوظائف تتكون من وحدتين فرعيتين، وحدة فرعية اقتطاعية (P08764) ووحدة فرعية مثيلة (P08763). تتعرف الوحدة الفرعية المثيلة على تسلسل الحمض النووي الخاص بالنظام وهي عبارة عن تعديل ناقلة الميثيل، على هذا النحو، فهو مكافئ وظيفيًا للوحدتين الفرعيتين M و S من نوكلياز الاقتطاع من النوع الأول. الوحدة الفرعية الاقتطاعية مطلوبة لاهتضام التقييد، على الرغم من أنه ليس له نشاط إنزيمي من تلقاء نفسه. تتعرف إنزيمات النوع الثالث على تسلسل الحمض النووي غير المتماثل الذي يتراوح طوله بين 5 و 6 زوج قاعدي وتقطع 25-27 زوج قاعدي عكس الاتجاه ومع الاتجاه لتترك نتوءات قصيرة مفردة 5'. إنها تتطلب وجود موقعين غير ميثليين مُوجَّهين بشكل عكسي للتعرف على عملية اهتضام التقييد. تعمل هذه الإنزيمات على مثيلة خيط واحد فقط من الحمض النووي، في موضع N-6 من بقايا الأدينوسيل، لذلك فإن الحمض النووي المتماثل حديثًا سيكون له خيط مثيلي واحد فقط، وهو ما يكفي للحماية من اهتضام التقييد. تنتمي إنزيمات النوع 3 إلى عائلة بيتا الفرعية من دنا ميتيل-ترانسفيراز، والتي تحتوي على العناصر التسعة التي تميز هذه العائلة، بما في ذلك العنصر 1، وجيب ربط S-أدينوسيل الميثيونين، والعنصر الرابع، المنطقة المحفزة (S/D/N (PP) Y/F). [36][37][28][38]
النوع 4
تتعرف إنزيمات النوع 4 على الحمض النووي المُمَثيَل المُعدَّل، ويُمثَّل بواسطة أنظمة McrBC و Mrr للإشريكية القولونية. [39]
النوع 5
تستخدم إنزيمات الاقتطاع من النوع الخامس (على سبيل المثال، مركب البروتين المرتبط بكريسبر 9-حمض نووي ريبوزي الإردشادي من كريسبر) الحمض النووي الريبوزي الإردشادي لاستهداف تسلسلات مُحددة غير متناظرة موجودة في الكائنات الغازية. يمكنهم قطع الحمض النووي بأطوال متغيرة، بشرط توفير الحمض النووي الريبوزي الإردشادي المناسب. إن مرونة هذه الإنزيمات وسهولة استخدامها تجعلها واعدة لتطبيقات الهندسة الوراثية المستقبلية. [39][40]
إنزيمات الاقتطاع الاصطناعية
يمكن إنشاء إنزيمات الاقتطاع الاصطناعية عن طريق دمج مجال ربط الحمض النووي الطبيعي أو الهندسي بمجال نوكلياز (غالبًا مجال الانقسام لنوع إنزيم اقتطاع من النوع 2 س Fok1).[41] يمكن أن تستهدف إنزيمات الاقتطاع الاصطناعية مواقع الحمض النووي الكبيرة (حتى 36 زوج قاعدي) ويمكن هندستها لربط تسلسلات الحمض النووي المرغوبة.[42] نوكلياز إصبع الزنك هي أكثر إنزيمات الاقتطاع الاصطناعية شيوعًا وتُستخدم بشكل عام في تطبيقات الهندسة الوراثية،[43][44][45][46] ولكن يمكن أيضًا استخدامها لمزيد من تطبيقات الاستنساخ الجيني القياسية.[47] تعتمد إنزيمات الاقتطاع الاصطناعية الأخرى على مجال ربط الحمض النووي لمؤثرات TAL. [48][49]
في عام 2013، صُمِّمَت تقنية "كريسبر-البروتين المرتبط بكريسبر 9" الجديدة، القائمة على نظام الدفاع الفيروسي بدائية النواة، لتعديل الجينوم، وسرعان ما حصل تبنّيها في المختبرات. [50]
يتم أيضًا تطوير الريبونوكليازات الاصطناعية التي تعمل بمثابة إنزيمات تقييدية للحمض النووي الريبوزي. نظام قائم على حمض نووي ببتيد (PNA)، يُسمى أزيمات الحمض النووي ببتيد (PNAzymes)، لديه مجموعة أكسيد النحاس الثنائي-2,9-ثنائي ميثيل فينانثرولين (Cu(II)-2,9-dimethylphenanthroline) التي تحاكي الريبونوكلياز لتسلسل معين من الحمض النووي الريبوزي وينقسم في منطقة غير ثنائية القاعدة (نتوء الحمض النووي الريبوزي) من مكون الحمض النووي الريبوزي المُستهدَف عندما يرتبط الإنزيم بالحمض النووي الريبوزي. يُظهِر هذا الإنزيم انتقائية من خلال القطع في موقع واحد فقط يكون إما لا يحتوي على عدم تطابق أو يُفضِّل حركيًا من موقعين مُحتمَلين للقطع. [51]
إنزيم القص النادر
إنزيم القص النادر (Rare-cutter enzyme)] هو إنزيم من إنزيمات الاقتطاع بمواقع تعرف تسلسل التعرف حيث يحدث بصورة نادرة في الجينوم. مثال على ذلك إنزيم NotI حيث يقوم بالقص بعد أول GC من التسلسل 5'-GCGGCCGC-3' ؛ إنزيمات التحديد بسبعة وثمانية أزواج من قواعد مواقع التعرف تسمى أيضا بإنزيمات القص النادر.
على سبيل المثال إنزيم القص النادر بسبعة مواقع تعرف نيوكليوتيدية يقص مرة واحدة كل 47 bp (16,384) bp التي تحتوي على ثمانية مواقع تعرف نيوكليوتيدية تقص كل 48 bp (65,536) bp على التوالي. وهي تستخدم في تسلسل الحمض النووي لقص الكروموسوم إلى قطع من هذه الإحجام.
انظر أيضًا
مراجع
- "معلومات عن إنزيمات الاقتطاع على موقع universalis.fr". universalis.fr. مؤرشف من الأصل في 25 يوليو 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "معلومات عن إنزيمات الاقتطاع على موقع id.loc.gov". id.loc.gov. مؤرشف من الأصل في 10 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "معلومات عن إنزيمات الاقتطاع على موقع britannica.com". britannica.com. مؤرشف من الأصل في 27 أبريل 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ernst L. (1987). From genes to clones : introduction to gene technology. Weinheim, Federal Republic of Germany ; New York, NY, USA : VCH. ISBN 978-0-89573-614-7. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bertani, G.; Weigle, J. J. (1953-02). "Host controlled variation in bacterial viruses". Journal of Bacteriology. 65 (2): 113–121. doi:10.1128/JB.65.2.113-121.1953. ISSN 0021-9193. PMID 13034700. مؤرشف من الأصل في 21 أغسطس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Arber, W.; Linn, S. (1969). "DNA modification and restriction". Annual Review of Biochemistry. 38: 467–500. doi:10.1146/annurev.bi.38.070169.002343. ISSN 0066-4154. PMID 4897066. مؤرشف من الأصل في 22 نوفمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Meselson, Matthew; Yuan, Robert (1968-03). "DNA Restriction Enzyme from E. coli". Nature (باللغة الإنجليزية). 217 (5134): 1110–1114. doi:10.1038/2171110a0. ISSN 1476-4687. مؤرشف من الأصل في 25 أبريل 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - "Host specificity of DNA produced by Escherichia coli. II. Control over acceptance of DNA from infecting phage lambda". مؤرشف من الأصل في 21 أغسطس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "DEGRADATION OF NON-REPLICATING BACTERIOPHAGE DNA IN NON-ACCEPTING CELLS". مؤرشف من الأصل في 21 أغسطس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Luria, S. E.; Human, Mary L. (1952-10). "A NONHEREDITARY, HOST-INDUCED VARIATION OF BACTERIAL VIRUSES1". Journal of Bacteriology. 64 (4): 557–569. ISSN 0021-9193. PMID 12999684. مؤرشف من الأصل في 25 أبريل 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - "How restriction enzymes became the workhorses of molecular biology". مؤرشف من الأصل في 14 نوفمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "A restriction enzyme from Hemophilus influenzae. I. Purification and general properties". مؤرشف من الأصل في 17 أكتوبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "A restriction enzyme from Hemophilus influenzae. II". مؤرشف من الأصل في 17 أكتوبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Loenen, Wil A. M.; Dryden, David T. F.; Raleigh, Elisabeth A.; Wilson, Geoffrey G.; Murray, Noreen E. (2014-01-01). "Highlights of the DNA cutters: a short history of the restriction enzymes". Nucleic Acids Research (باللغة الإنجليزية). 42 (1): 3–19. doi:10.1093/nar/gkt990. ISSN 0305-1048. مؤرشف من الأصل في 24 يونيو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Specific cleavage of simian virus 40 DNA by restriction endonuclease of Hemophilus influenzae". مؤرشف من الأصل في 22 نوفمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The Nobel Prize in Physiology or Medicine 1978". NobelPrize.org (باللغة الإنجليزية). مؤرشف من الأصل في 08 فبراير 2021. اطلع عليه بتاريخ 09 فبراير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "A bacterial clone synthesizing proinsulin". مؤرشف من الأصل في 5 فبراير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Restriction Endonucleases. Berlin, Heidelberg: Springer Berlin Heidelberg. 2004. ISBN 978-3-642-18851-0. OCLC 840291580. مؤرشف من الأصل في 10 فبراير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Smith, Hamilton O.; Nathans, Daniel (1973-12-15). "A Suggested nomenclature for bacterial host modification and restriction systems and their enzymes". Journal of Molecular Biology (باللغة الإنجليزية). 81 (3): 419–423. doi:10.1016/0022-2836(73)90152-6. ISSN 0022-2836. مؤرشف من الأصل في 10 فبراير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Roberts, Richard J.; Belfort, Marlene; Bestor, Timothy; Bhagwat, Ashok S.; Bickle, Thomas A.; Bitinaite, Jurate; Blumenthal, Robert M.; Degtyarev, Sergey Kh; Dryden, David T. F. (2003-04-01). "A nomenclature for restriction enzymes, DNA methyltransferases, homing endonucleases and their genes". Nucleic Acids Research (باللغة الإنجليزية). 31 (7): 1805–1812. doi:10.1093/nar/gkg274. ISSN 0305-1048. مؤرشف من الأصل في 28 أغسطس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Jeltsch, Albert; Kröger, Manfred; Pingoud, Alfred (1995-07-04). "Evidence for an evolutionary relationship among type-II restriction endonucleases". Gene (باللغة الإنجليزية). 160 (1): 7–16. doi:10.1016/0378-1119(95)00181-5. ISSN 0378-1119. مؤرشف من الأصل في 9 فبراير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Jeltsch, Albert; Pingoud, Alfred (1996-02-01). "Horizontal gene transfer contributes to the wide distribution and evolution of type II restriction-modification systems". Journal of Molecular Evolution (باللغة الإنجليزية). 42 (2): 91–96. doi:10.1007/BF02198833. ISSN 1432-1432. مؤرشف من الأصل في 13 يونيو 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Naito, T.; Kusano, K.; Kobayashi, I. (1995-02-10). "Selfish behavior of restriction-modification systems". Science (باللغة الإنجليزية). 267 (5199): 897–899. doi:10.1126/science.7846533. ISSN 0036-8075. PMID 7846533. مؤرشف من الأصل في 24 يونيو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bickle, T. A.; Krüger, D. H. (1993-06). "Biology of DNA restriction". Microbiological Reviews. 57 (2): 434–450. ISSN 0146-0749. PMID 8336674. مؤرشف من الأصل في 13 يناير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Boyer, H. W. (1971). "DNA restriction and modification mechanisms in bacteria". Annual Review of Microbiology. 25: 153–176. doi:10.1146/annurev.mi.25.100171.001101. ISSN 0066-4227. PMID 4949033. مؤرشف من الأصل في 22 نوفمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Yuan, R. (1981). "Structure and mechanism of multifunctional restriction endonucleases". Annual Review of Biochemistry. 50: 285–319. doi:10.1146/annurev.bi.50.070181.001441. ISSN 0066-4154. PMID 6267988. مؤرشف من الأصل في 17 أكتوبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Types of Restriction Endonucleases | NEB". www.neb.com. مؤرشف من الأصل في 24 ديسمبر 2020. اطلع عليه بتاريخ 09 فبراير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Sistla, Srivani; Rao, Desirazu N. (2004-01). "S-Adenosyl-L-methionine-dependent restriction enzymes". Critical Reviews in Biochemistry and Molecular Biology. 39 (1): 1–19. doi:10.1080/10409230490440532. ISSN 1040-9238. PMID 15121719. مؤرشف من الأصل في 21 أغسطس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Williams, Raymond J. (2003-03). "Restriction endonucleases: classification, properties, and applications". Molecular Biotechnology. 23 (3): 225–243. doi:10.1385/mb:23:3:225. ISSN 1073-6085. PMID 12665693. مؤرشف من الأصل في 20 أغسطس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Murray, N. E. (2000-06). "Type I restriction systems: sophisticated molecular machines (a legacy of Bertani and Weigle)". Microbiology and molecular biology reviews: MMBR. 64 (2): 412–434. doi:10.1128/mmbr.64.2.412-434.2000. ISSN 1092-2172. PMID 10839821. مؤرشف من الأصل في 20 أغسطس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Restriction endonucleases. Berlin: Springer-Verlag. 2004. ISBN 3-540-20502-0. OCLC 53476933. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Fundamental laboratory approaches for biochemistry and biotechnology (الطبعة 2nd ed). Hoboken, NJ: John Wiley. 2010. ISBN 978-0-470-08766-4. OCLC 420027217. مؤرشف من الأصل في 13 أبريل 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة)صيانة CS1: نص إضافي (link) - Siwek, Wojciech; Czapinska, Honorata; Bochtler, Matthias; Bujnicki, Janusz M.; Skowronek, Krzysztof (2012-08). "Crystal structure and mechanism of action of the N6-methyladenine-dependent type IIM restriction endonuclease R.DpnI". Nucleic Acids Research. 40 (15): 7563–7572. doi:10.1093/nar/gks428. ISSN 1362-4962. PMID 22610857. مؤرشف من الأصل في 04 نوفمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Mierzejewska, Karolina; Siwek, Wojciech; Czapinska, Honorata; Kaus-Drobek, Magdalena; Radlinska, Monika; Skowronek, Krzysztof; Bujnicki, Janusz M.; Dadlez, Michal; Bochtler, Matthias (2014-07). "Structural basis of the methylation specificity of R.DpnI". Nucleic Acids Research. 42 (13): 8745–8754. doi:10.1093/nar/gku546. ISSN 1362-4962. PMID 24966351. مؤرشف من الأصل في 03 سبتمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Pingoud, A.; Jeltsch, A. (2001-09-15). "Structure and function of type II restriction endonucleases". Nucleic Acids Research. 29 (18): 3705–3727. doi:10.1093/nar/29.18.3705. ISSN 1362-4962. PMID 11557805. مؤرشف من الأصل في 20 أغسطس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Dryden, D. T.; Murray, N. E.; Rao, D. N. (2001-09-15). "Nucleoside triphosphate-dependent restriction enzymes". Nucleic Acids Research. 29 (18): 3728–3741. doi:10.1093/nar/29.18.3728. ISSN 1362-4962. PMID 11557806. مؤرشف من الأصل في 20 أغسطس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Meisel, A.; Bickle, T. A.; Krüger, D. H.; Schroeder, C. (1992-01-30). "Type III restriction enzymes need two inversely oriented recognition sites for DNA cleavage". Nature. 355 (6359): 467–469. doi:10.1038/355467a0. ISSN 0028-0836. PMID 1734285. مؤرشف من الأصل في 28 أغسطس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bourniquel, Aude A.; Bickle, Thomas A. (2002-11). "Complex restriction enzymes: NTP-driven molecular motors". Biochimie. 84 (11): 1047–1059. doi:10.1016/s0300-9084(02)00020-2. ISSN 0300-9084. PMID 12595133. مؤرشف من الأصل في 20 أغسطس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - "Types of Restriction Endonucleases | NEB". www.neb.com. مؤرشف من الأصل في 24 ديسمبر 2020. اطلع عليه بتاريخ 10 فبراير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Horvath, Philippe; Barrangou, Rodolphe (2010-01-08). "CRISPR/Cas, the immune system of bacteria and archaea". Science (New York, N.Y.). 327 (5962): 167–170. doi:10.1126/science.1179555. ISSN 1095-9203. PMID 20056882. مؤرشف من الأصل في 14 ديسمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Hybrid restriction enzymes: zinc finger fusions to Fok I cleavage domain". مؤرشف من الأصل في 26 نوفمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Urnov, Fyodor D.; Rebar, Edward J.; Holmes, Michael C.; Zhang, H. Steve; Gregory, Philip D. (2010-09). "Genome editing with engineered zinc finger nucleases". Nature Reviews Genetics (باللغة الإنجليزية). 11 (9): 636–646. doi:10.1038/nrg2842. ISSN 1471-0064. مؤرشف من الأصل في 06 نوفمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Townsend, Jeffrey A.; Wright, David A.; Winfrey, Ronnie J.; Fu, Fengli; Maeder, Morgan L.; Joung, J. Keith; Voytas, Daniel F. (2009-05). "High-frequency modification of plant genes using engineered zinc-finger nucleases". Nature (باللغة الإنجليزية). 459 (7245): 442–445. doi:10.1038/nature07845. ISSN 1476-4687. مؤرشف من الأصل في 10 فبراير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Shukla, Vipula K.; Doyon, Yannick; Miller, Jeffrey C.; DeKelver, Russell C.; Moehle, Erica A.; Worden, Sarah E.; Mitchell, Jon C.; Arnold, Nicole L.; Gopalan, Sunita (2009-05). "Precise genome modification in the crop species Zea mays using zinc-finger nucleases". Nature (باللغة الإنجليزية). 459 (7245): 437–441. doi:10.1038/nature07992. ISSN 1476-4687. مؤرشف من الأصل في 5 مايو 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ=(مساعدة) - Ekker, Stephen C. (2008). "Zinc finger-based knockout punches for zebrafish genes". Zebrafish. 5 (2): 121–123. doi:10.1089/zeb.2008.9988. ISSN 1557-8542. PMID 18554175. مؤرشف من الأصل في 15 نوفمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Geurts, Aron M.; Cost, Gregory J.; Freyvert, Yevgeniy; Zeitler, Bryan; Miller, Jeffrey C.; Choi, Vivian M.; Jenkins, Shirin S.; Wood, Adam; Cui, Xiaoxia (2009-07-24). "Knockout rats via embryo microinjection of zinc-finger nucleases". Science (New York, N.Y.). 325 (5939): 433. doi:10.1126/science.1172447. ISSN 1095-9203. PMID 19628861. مؤرشف من الأصل في 16 نوفمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Tovkach, Andriy; Zeevi, Vardit; Tzfira, Tzvi (2011-01-10). "Expression, purification and characterization of cloning-grade zinc finger nuclease". Journal of Biotechnology (باللغة الإنجليزية). 151 (1): 1–8. doi:10.1016/j.jbiotec.2010.10.071. ISSN 0168-1656. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Christian, Michelle; Cermak, Tomas; Doyle, Erin L.; Schmidt, Clarice; Zhang, Feng; Hummel, Aaron; Bogdanove, Adam J.; Voytas, Daniel F. (2010-10-01). "Targeting DNA Double-Strand Breaks with TAL Effector Nucleases". Genetics (باللغة الإنجليزية). 186 (2): 757–761. doi:10.1534/genetics.110.120717. ISSN 0016-6731. PMID 20660643. مؤرشف من الأصل في 26 يناير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Li, Ting; Huang, Sheng; Jiang, Wen Zhi; Wright, David; Spalding, Martin H.; Weeks, Donald P.; Yang, Bing (2011-01-01). "TAL nucleases (TALNs): hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain". Nucleic Acids Research (باللغة الإنجليزية). 39 (1): 359–372. doi:10.1093/nar/gkq704. ISSN 0305-1048. مؤرشف من الأصل في 12 أكتوبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Hsu, Patrick D.; Lander, Eric S.; Zhang, Feng (2014-06-05). "Development and applications of CRISPR-Cas9 for genome engineering". Cell. 157 (6): 1262–1278. doi:10.1016/j.cell.2014.05.010. ISSN 1097-4172. PMID 24906146. مؤرشف من الأصل في 22 يناير 2021. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "PNAzymes that are artificial RNA restriction enzymes". مؤرشف من الأصل في 16 نوفمبر 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة)
- "الموسوعة العربية :: ENCYCLOPEDIA". مؤرشف من الأصل في 28 أبريل 2016. اطلع عليه بتاريخ 23 يونيو 2014. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "الإنزيمات القاطعة Restriction Nucleases". مؤرشف من الأصل في 05 يونيو 2019. اطلع عليه بتاريخ 23 يونيو 2014. الوسيط
|CitationClass=تم تجاهله (مساعدة) - نسخة مؤرشفة
 بوابة علم الأحياء
بوابة علم الأحياء بوابة تقانة
بوابة تقانة بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة تقانة حيوية
بوابة تقانة حيوية بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي