نقل محواري
النقل المحواري[1] ويُسمى أيضًا نقل أو جريان هيولى المحور[2] (بالإنجليزية: Axonal transport) هو عملية خلوية مسئولة عن حركة الميتوكوندريا والدهون والحويصلات المشبكية والبروتينات والعضيات الأخرى من وإلى جسم الخلايا العصبية من خلال سيتوبلازم محور الليفة العصبية المسمى بـ سيتوبلازم المحور العصبي[3]، بالرغم من أن بعض المحاور العصبية يصل طولها إلى عدة أمتار إلا أن الخلايا العصبية لا يمكنها الاعتماد على ظاهرة الانتشار لحمل منتجات النواة والعضيات إلى نهاية محورها العصبي، النقل المحوري العصبي مسئول أيضًا عن نقل الجزيئات -التي يتم تكسيرها- من محور الليفة العصبية رجوعًا إلى جسم الخلية، حيث يتم تكسيرها بواسطة اللايسوسومات.[4]
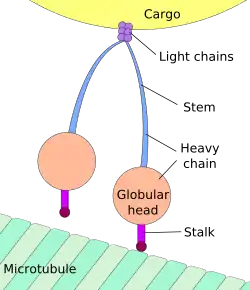
الحركة في اتجاه جسم الخلية تسمى بالنقل الرجوعي، بينما الحركة في اتجاه التشابك العصبي تسمى بالنقل التقدمي.[5][6]
آلية العمل
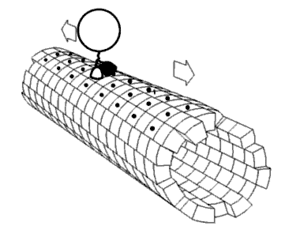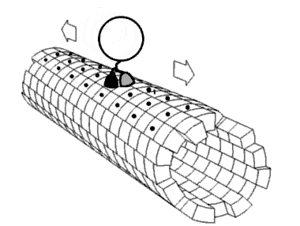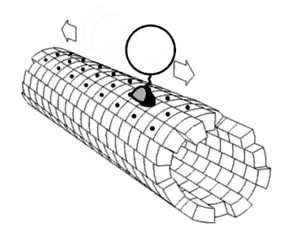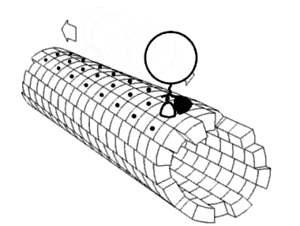
يتم تصنيع الأغلبية العظمى من بروتينات محور الليفة العصبية داخل جسم الخلية العصبية ويتم نقلها بطول المحاور العصبية، لوحظ حدوث عملية ترجمة لبعض جزيئات mRNA داخل المحاور العصبية[7][8]، يحدث النقل المحوري العصبي أثناء حياة الخلية العصبية ويعد أساسيًا لنموها وبقائها، تجري الأنيبيبات الدقيقة (المصنوعة من تيوبيولين) على طول المحور العصبي وتعطي المسارات الهيكلية الخلوية الرئيسية للنقل، تعتبر الكينيسين والداينين هي البروتينات الحركية التي تحرك المركبات في الاتجاه التقدمي (من جسم الخلية العصبية إلى طرف المحور العصبي) والاتجاه الرجوعي (عائدة إلى جسم الخلية العصبية) على التوالي، ترتبط البروتينات الحركية ببعضها وتقوم بنقل العديد من المركبات المختلفة بما في ذلك الميتوكوندريا و بوليميرات هيكل الخلية والجسيمات البلعمية الذاتية والحويصلات المشبكية المحتوية على النواقل العصبية. قد يكون النقل المحوري العصبي سريعًا أوبطئيًا، كما أنه قد يكون في اتجاه تقدمي (بعيدًا عن جسم الخلية العصبية) أو في اتجاه رجوعي (ينقل المواد من المحور العصبي إلى جسم الخلية).
النقل السريع والبطيء
تتحرك المركبات الحويصلية بسرعة 50-400 ميللي متر/اليوم، بينما تأخذ البروتينات القابلة للذوبان (العصارية الخلوية) والبروتينات الهيكلية الخلوية وقتًا أطول حيث تتحرك بسرعة أقل من 8ميللي متر/اليوم.
لقد تم تفسير الآلية الأساسية للنقل المحوري العصبي السريع منذ عقود، ولكن النقل المحوري العصبي البطيء أصبح واضحًا في الآونة الأخيرة فقط كنتيجة تقنيات التصوير المتقدمة[9]، وقد مكنت تقنيات وضع العلامات الفلورية (مثل: المجهر الفلوري) الرؤية المباشرة للنقل المحوري العصبي في الخلية العصبية الحية.
وقد كشفت الدراسات الحديثة أن حركة المركبات الهيكلية الخلوية البطيئة تعد في الأصل سريعة ولكن على عكس المركبات السريعة فهي متكررة الوقوف مما يجعل معدل النقل الإجمالي أبطأ كثيرًا. تعرف آلية العمل هذه بنموذج "توقف وانطلق" للنقل المحوري البطيء، وقد تم التحقق منها على نطاق واسع لنقل الشعيرات العصبية للبروتينات الهيكلية.[10] تعد حركة المركبات القابلة للذوبان (العصارية الخلوية) أكثر تعقيدًا، ولكن يبدو أن لها أساسًا متشابهًا مع تجمع البروتينات القابلة للذوبان في صورة مركبات متعددة البروتين والتي يتم نقلها بواسطة تفاعلات مؤقتة مع المركبات الأكثر سرعة أثناء حركتها في النقل المحوري السريع.[11][12][13]
الأمر أشبه بالفرق في معدلات النقل بين قطارات مترو الأنفاق المحلية والسريعة، فعلى الرغم من أن كلا النوعين من القطارات يسافران بسرعات متشابهة بين المحطات فإن القطار المحلي يستغرق وقتًا أطول بكثير للوصول إلى نهاية الخط لأنه يتوقف في كل محطة في حين أن القطار السريع لا يتوقف إلا على بعض محطات الطريق.
النقل التقدمي
يعرف النقل التقدمي -والمسمى أيضا بـ "النقل منتصب المشية"- بحركة الجزيئات والعضيات إلى الخارج أي من جسم الخلية إلى منطقة التشابك العصبي أوغشاء الخلية.
تتم الحركة التقدمية للمركبات الفردية في حويصلات النقل لكل من المكونات السريعة والبطيئة على طول الأنيبيبات الدقيقة[6] بواسطة بروتينات الكينيسين.[4] بالرغم من أن العديد من بروتينات الكينيسين مسئولة عن النقل البطيء[9] فإن آلية التوقُّف أثناء نقل المركبات البطيئة لا تزال مجهولة.
هناك فئتان من النقل التقدمي البطيء: المكون البطيء أ (SCa) الذي يحمل بشكل رئيسي الأنيبيبات الدقيقة والخيوط العصبية بمعدل 1-0.1 ميللي متر/اليوم، والمكون البطيء ب (SCb) الذي يحمل أكثر من 200 نوع من البروتينات المتنوعة والأكتين بمعدل قد يصل إلى 6 ميللي متر/اليوم.[9] يتم نقل المكون البطيء b والذي يحمل أيضًا الأكتين بمعدل 2-3 ميللي متر /اليوم في محاور الخلية الشبكية.
أثناء إعادة التنشيط من الكمون يدخل فيروس الهربس البسيط (HSC) في دورة تحلله، ويستخدم آليات النقل التقدمي في الانتقال من الخلايا العصبية للعقد الجزرية الظهرية إلى الجلد أو الغشاء المخاطي الذي سيصيبه لاحقًا.[14] لقد عرف مستقبل المركبات لمحركات للنقل التقدمي -بروتين الكينيسين- على أنه بروتين الطليعة النشواني (APP) وهو البروتين الرئيسي الذي ينتج لويحات الشيخوخة الموجودة في مرض ألزهايمر،[15] يرتبط ببتيد مكون من 15 حمض أميني موجود في نهاية الكربوكسيل السيتوبلازمي لل APP بقوة مع الكينيسين التقليدي kinesin-1 ويقوم بنقل المركبات الخارجية في المحور العصبي العملاق.[16]
يتحرك المنغنيز -وهو عامل التباين للتصوير بالرنين المغناطيسي T1-MRI- بالنقل التقدمي بعد إجراء الحقن التجسيمي في دماغ حيوانات التجارب والبتالي يقوم بكشف الدوائر عن طريق التصوير بالرنين المغناطيسي للدماغ في الحيوانات الحية كما كانت هذه العملية رائدة من قبل روبيا باتلر و إلين بيرر و روس جاكوبس.
وكشفت الدراسات التي أجريت على الفئران على السلسة الخفيفة kinesin-1 أن عنصر المنغنيز Mn يتحرك بواسطة النقل القائم على الكينيسين في العصب البصري وفي الدماغ.
يعتمد النقل في كل من النتوءات الحصينية والعصب البصري كذلك على بروتين الطليعة النشواني[17]، ويقل معدل النقل من النتوءات الحصينية إلى الدماغ الأمامي عند الشيخوخة ويتم تغيير الوجهة من خلال وجود لويحات مرض ألزهايمر.[18]
النقل الرجوعي
يقوم النقل الرجوعي بنقل الجزيئات/العضيات بعيدًا عن نهايات المحور العصبي نحو جسم الخلية، يتم النقل المحوري الرجوعي بواسطة الداينين السيتوبلازمي، ويستخدم على سبيل المثال لإرسال رسائل كيميائية ومنتجات الإدخال الخلوي المتجهة إلى الليسوسومات من المحور العصبي رجوعًا إلى الخلية[4]، وبالعمل بمتوسط سرعة 2 ميكرومتر في الثانية داخل الجسم الحي[19][20] فإنه يمكن أن يغطي النقل الرجعي السريع 10-20 سنتي متر في اليوم،[4] يقوم النقل الرجوعي السريع بإعادة الحويصلات المشبكية المستخدمة والمواد الأخرى إلى جسم الخلية، ويقوم بإبلاغ جسم الخلية بالظروف الموجودة في النهايات العصبية للمحور، يحمل النقل الرجوعي الإشارات اللازمة للبقاء من التشابك العصبي رجوعًا إلى جسم الخلية مثل مستقبل عامل نمو الأعصاب (TRK).[21]
تستغل بعض مسببات الأمراض هذه العملية لغزو الجهاز العصبي، فهي تدخل أطراف المحور العصبي وتتوجه إلى جسم الخلية بواسطة النقل الرجوعي، وتشمل الأمثلة المادة السامة للكزاز (التيتانوس) وفيروسات الهربس البسيط والسعار وشلل الأطفال، في مثل هذه العدوى يرجع التأخير بين العدوى وظهور الأعراض إلى الوقت اللازم لوصول مسببات الأمراض إلى جسم الخلية.[22] يتحرك فيروس الهربس البسيط في الاتجاهين في المحاور العصبية اعتمادًا على دورة حياته، فالنقل الرجوعي يسيطر على قطبية القفيصات الواردة.[23]
عواقب الانقطاع
عندما يتم إعاقة أو إيقاف النقل المحوري يتحول علم وظائف الأعضاء الطبيعي إلى الفيزيولوجيا المرضية وربما يحدث تجمع لسيتوبلازم المحور العصبي المسمى بالجسم الكروي المحواري، نظرًا لأن النقل المحوري يمكن أن يتم تعطيله بعدة طرق فإنه يمكن رؤية الأجسام الكروية للمحور في العديد من فئات الأمراض المختلفة بما في ذلك الأمراض الوراثية والصدمية والإقفارية والمعدية والسامة والتنكسية والخاصة بالمادة البيضاء التي تسمى اعتلال بيضاء الدماغ.
ترتبط العديد من الأمراض التنكسية العصبية النادرة بالطفرات الجينية في البروتينات الحركية الكينيسين والدينين، وفي هذه الحالات من المرجح أن يكون النقل المحواري هو السبب الرئيسي في إحداث المرض.[24]
يرتبط النقل المحواري المختل أيضًا بأشكال متفرقة (شائعة) من أمراض الأعصاب مثل آلزهايمر وباركنسون (الشلل الرعاش)[9]، ويرجع ذلك بشكل أساسي إلى وجود ملاحظات عديدة أن هناك تراكمات كبيرة في المحور العصبي تظهر دائمًا في الخلايا العصبية المصابة، وهذه الجينات تلعب دورًا في الأشكال العائلية لهذه الأمراض ولديها أيضًا أدوار مزعومة في النقل المحوري العادي، ومع ذلك فإن هناك القليل من الأدلة المباشرة على دور النقل المحوري في إحداث الأمراض الأخيرة، وقد تكون الآليات الأخرى مثل السمية العصبية المباشرة أكثر صلة.
يؤدي توقف تدفق سيتوبلازم المحور على حافة المنطقة الإقفارية في اعتلال الشبكية الوعائي إلى تورم الألياف العصبية التي تؤدي إلى نضحات ناعمة أو بقع صوفية قطنية.
نظرًا لأن المحور العصبي يعتمد على النقل المحوري للبروتينات والمواد الحيوية، فإن الإصابة مثل إصابة المحور العصبي المنتشرة والتي تعيق النقل ستتسبب في تدهور المحور العصبي البعيد في عملية تسمى تنكس واليريان، يمكن أن تسبب عقاقير السرطان التي تعطل النمو السرطاني عن طريق تغيير الأنيبيبات الدقيقة -الضرورية لانقسام الخلايا)- في إتلاف الأعصاب لأن الأنيبيبات الدقيقة ضرورية للنقل المحوري.
العدوى
يصل فيروس داء الكلب (ريبس) إلى الجهاز العصبي المركزي عن طريق النقل المحوري الرجوعي[25]، يتم إدخال السم العصبي الكزازي في الموصل العصبي العضلي من خلال ربط بروتينات النيدوجين، ويتم نقله إلى الوراء نحو جسم الخلية العصبية أثناء الإشارة إلى الدخلولات[26]، تنتقل الفيروسات الموجهة للعصب -مثل فيروس الهربس- داخل المحاور العصبية باستخادم آلات النقل الخلوي كما هو موضح في عمل مجموعة إلين بيرر[27][28]، من المتوقع أن يكون هناك بعض العوامل المعدية الأخرى التي تستخدم النقل المحوري.[29] يعتقد الآن أن مثل هذه العدوى تساهم في مرض آلزهايمر وغيره من الاضطرابات العصبية التنكسية.[30][31]
المراجع
- "Al-Qamoos القاموس | English Arabic dictionary / قاموس إنجليزي عربي". www.alqamoos.org. مؤرشف من الأصل في 11 يوليو 2020. اطلع عليه بتاريخ 11 يوليو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Al-Qamoos القاموس | English Arabic dictionary / قاموس إنجليزي عربي". www.alqamoos.org. مؤرشف من الأصل في 11 يوليو 2020. اطلع عليه بتاريخ 11 يوليو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Sabry J, O'Connor TP, Kirschner MW (June 1995). "Axonal transport of tubulin in Ti1 pioneer neurons in situ". Neuron. 14 (6): 1247–56. doi:10.1016/0896-6273(95)90271-6. PMID 7541635. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Oztas E (2003). "Neuronal Tracing" (PDF). Neuroanatomy. 2: 2–5. مؤرشف من الأصل (PDF) في 25 أكتوبر 2005. اطلع عليه بتاريخ 10 يوليو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Karp G, van der Geer P. Cell and molecular biology: concepts and experiments (الطبعة 4th). John Wiley. صفحة 344. ISBN 978-0-471-46580-5. مؤرشف من الأصل في 10 يوليو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bear MF, Connors BW, Paradso MA (2007). Neuroscience : exploring the brain (الطبعة 3rd). Lippincott Williams & Wilkins. صفحة 41. ISBN 978-0-7817-6003-4. مؤرشف من الأصل في 10 يوليو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Giustetto M, Hegde AN, Si K, Casadio A, Inokuchi K, Pei W, Kandel ER, Schwartz JH (November 2003). "Axonal transport of eukaryotic translation elongation factor 1alpha mRNA couples transcription in the nucleus to long-term facilitation at the synapse". Proceedings of the National Academy of Sciences of the United States of America. 100 (23): 13680–5. doi:10.1073/pnas.1835674100. PMC 263873. PMID 14578450. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Si K, Giustetto Si K, Giustetto M, Etkin A, Hsu R, Janisiewicz AM, Miniaci MC, Kim JH, Zhu H, Kandel ER (December 2003). "A neuronal isoform of CPEB regulates local protein synthesis and stabilizes synapse-specific long-term facilitation in aplysia". Cell. 115 (7): 893–904. doi:10.1016/s0092-8674(03)01021-3. PMID 14697206. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Roy S, Zhang B, Lee VM, Trojanowski JQ (January 2005). "Axonal transport defects: a common theme in neurodegenerative diseases". Acta Neuropathologica. 109 (1): 5–13. doi:10.1007/s00401-004-0952-x. PMID 15645263. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Brown A (March 2003). "Axonal transport of membranous and nonmembranous cargoes: a unified perspective". The Journal of Cell Biology. 160 (6): 817–21. doi:10.1083/jcb.200212017. PMC 2173776. PMID 12642609. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Scott DA, Das U, Tang Y, Roy S (May 2011). "Mechanistic logic underlying the axonal transport of cytosolic proteins". Neuron. 70 (3): 441–54. doi:10.1016/j.neuron.2011.03.022. PMC 3096075. PMID 21555071. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Roy S, Winton MJ, Black MM, Trojanowski JQ, Lee VM (March 2007). "Rapid and intermittent cotransport of slow component-b proteins". The Journal of Neuroscience. 27 (12): 3131–8. doi:10.1523/JNEUROSCI.4999-06.2007. PMC 6672457. PMID 17376974. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Kuznetsov AV (2011). "Analytical solution of equations describing slow axonal transport based on the stop-and-go hypothesis". Central European Journal of Physics. 9 (3): 662–673. doi:10.2478/s11534-010-0066-0. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Holland DJ, Miranda-Saksena M, Boadle RA, Armati P, Cunningham AL (October 1999). "Anterograde transport of herpes simplex virus proteins in axons of peripheral human fetal neurons: an immunoelectron microscopy study". Journal of Virology. 73 (10): 8503–11. PMC 112870. PMID 10482603. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Satpute-Krishnan P, DeGiorgis JA, Conley MP, Jang M, Bearer EL (October 2006). "A peptide zipcode sufficient for anterograde transport within amyloid precursor protein". Proceedings of the National Academy of Sciences of the United States of America. 103 (44): 16532–7. doi:10.1073/pnas.0607527103. PMC 1621108. PMID 17062754. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Seamster PE, Loewenberg M, Pascal J, Chauviere A, Gonzales A, Cristini V, Bearer EL (October 2012). "Quantitative measurements and modeling of cargo-motor interactions during fast transport in the living axon". Physical Biology. 9 (5): 055005. doi:10.1088/1478-3975/9/5/055005. PMC 3625656. PMID 23011729. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Gallagher JJ, Zhang X, Ziomek GJ, Jacobs RE, Bearer EL (April 2012). "Deficits in axonal transport in hippocampal-based circuitry and the visual pathway in APP knock-out animals witnessed by manganese enhanced MRI". NeuroImage. 60 (3): 1856–66. doi:10.1016/j.neuroimage.2012.01.132. PMC 3328142. PMID 22500926. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bearer EL, Manifold-Wheeler BC, Medina CS, Gonzales AG, Chaves FL, Jacobs RE (October 2018). "Alterations of functional circuitry in aging brain and the impact of mutated APP expression". Neurobiology of Aging. 70: 276–290. doi:10.1016/j.neurobiolaging.2018.06.018. PMC 6159914. PMID 30055413. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Gibbs KL, Kalmar B, Sleigh JN, Greensmith L, Schiavo G (January 2016). "In vivo imaging of axonal transport in murine motor and sensory neurons". Journal of Neuroscience Methods. 257: 26–33. doi:10.1016/j.jneumeth.2015.09.018. PMC 4666412. PMID 26424507. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Sleigh J, Schiavo G (2016). "Older but not slower: aging does not alter axonal transport dynamics of signalling endosomes in vivo". Matters. 2 (6). doi:10.19185/matters.201605000018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cui B, Wu C, Chen L, Ramirez A, Bearer EL, Li WP, Mobley WC, Chu S (August 2007). "One at a time, live tracking of NGF axonal transport using quantum dots". Proceedings of the National Academy of Sciences of the United States of America. 104 (34): 13666–71. doi:10.1073/pnas.0706192104. PMC 1959439. PMID 17698956. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Saladin, Kenneth. Anatomy and Physiology: The Unity of Form and Function. Sixth. New York : McGraw-Hill, 2010. 445. Print.
- Bearer EL, Breakefield XO, Schuback D, Reese TS, LaVail JH (July 2000). "Retrograde axonal transport of herpes simplex virus: evidence for a single mechanism and a role for tegument". Proceedings of the National Academy of Sciences of the United States of America. 97 (14): 8146–50. doi:10.1073/pnas.97.14.8146. PMC 16684. PMID 10884436. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Maday S, Twelvetrees AE, Moughamian AJ, Holzbaur EL (October 2014). "Axonal transport: cargo-specific mechanisms of motility and regulation". Neuron. 84 (2): 292–309. doi:10.1016/j.neuron.2014.10.019. PMC 4269290. PMID 25374356. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Mitrabhakdi E, Shuangshoti S, Wannakrairot P, Lewis RA, Susuki K, Laothamatas J, Hemachudha T (November 2005). "Difference in neuropathogenetic mechanisms in human furious and paralytic rabies". Journal of the Neurological Sciences. 238 (1–2): 3–10. doi:10.1016/j.jns.2005.05.004. PMID 16226769. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bercsenyi K, Schmieg N, Bryson JB, Wallace M, Caccin P, Golding M, Zanotti G, Greensmith L, Nischt R, Schiavo G (November 2014). "Tetanus toxin entry. Nidogens are therapeutic targets for the prevention of tetanus" (PDF). Science. 346 (6213): 1118–23. doi:10.1126/science.1258138. PMID 25430769. مؤرشف (PDF) من الأصل في 22 ديسمبر 2016. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Satpute-Krishnan P, DeGiorgis JA, Bearer EL (December 2003). "Fast anterograde transport of herpes simplex virus: role for the amyloid precursor protein of alzheimer's disease". Aging Cell. 2 (6): 305–18. doi:10.1046/j.1474-9728.2003.00069.x. PMC 3622731. PMID 14677633. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cheng SB, Ferland P, Webster P, Bearer EL (March 2011). "Herpes simplex virus dances with amyloid precursor protein while exiting the cell". PLOS ONE. 6 (3): e17966. doi:10.1371/journal.pone.0017966. PMC 3069030. PMID 21483850. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bearer EL, Satpute-Krishnan P (September 2002). "The role of the cytoskeleton in the life cycle of viruses and intracellular bacteria: tracks, motors, and polymerization machines". Current Drug Targets. Infectious Disorders. 2 (3): 247–64. doi:10.2174/1568005023342407. PMC 3616324. PMID 12462128. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Itzhaki RF, Lathe R, Balin BJ, Ball MJ, Bearer EL, Braak H, et al. (2016). "Microbes and Alzheimer's Disease". Journal of Alzheimer's Disease. 51 (4): 979–84. doi:10.3233/JAD-160152. PMC 5457904. PMID 26967229. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "No place like asphalt for these hardy microbes". New Scientist. 206 (2757): 15. 2010. doi:10.1016/s0262-4079(10)60991-8. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة طب
بوابة طب