نشوء تعايشي
تنص نظرية النشوء التعايشي (الارتقاء) على أن هناك العديد من العضيات حقيقية النوى نشأت بين الكائنات الحية وحيدة الخلية وتصرفت على أنها خلية واحدة. ومن الامثلة الأكثر شيوعاً على هذه النظرية، هي المايتوكندريا والبلاستيدات الخضراء. تبين أن أصول بعض العضيات مثل المايتوكندريا والبلاستيدات الخضراء في الخلايا حقيقية النواة، وأن البلاستيدات الخضراء والتي قامت الخلايا حقيقة النوى ببلعمتها، وأن الخلايا والبكتيريا المحاصرين في داخلها دخلت علاقة تكافلية، هذا يعني أن واحدا من الكائنات الحية ( البكتيريا ) عاش داخل الخلايا حقيقية النواة الأخرى، وتصرفت على أنها خلية واحدة بحيث لا يمكن لخلية أن تعيش دون وجود الخلية الأخرى. السبب وراء النظر في أن الميتوكندريا والبلاستيدات الخضراء يعود أصلها إلى الخلايا بدائية النواة هو احتوائها على العديد من الانزيمات وانظمة النقل المماثلة لتلك الموجودة في البكتيريا.
اثباتات على النظرية
من حيث الشكل
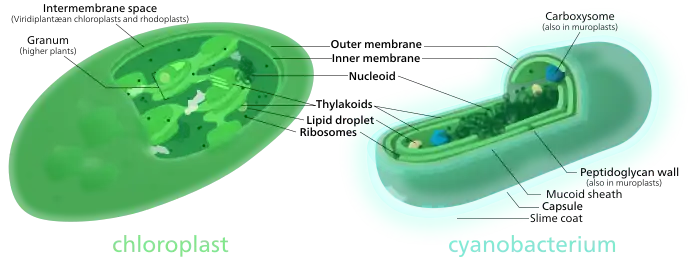
- حجم وشكل المايتوكندريا والبلاستيدات الخضراء مشابه لحجم وشكل الخلايا البكتيرية.[1]
- تمتلك المايتوكندريا والبلاستيدات الخضراء غشاء خلوي مزدوج كالخلايا العادية والبكتيرية.[1]
- البنية الداخلية والكيمياء الحيوية للبلاستيدات هي مشابهة جدا لتلك التي في البكتيريا الزرقاء، مثل وجود الثايلاكويدات وانواع معينة من صبغة الكلوروفيل. كما ان تقديرات النشوء والتطور التي شيدت مع البكتيريا، البلاستيدات وجينومات حقيقة النوى، تشير إلى ان البلاستيدات هي الأكثر ارتباطها بالبكتيريا الزرقاء.
من حيث التركيب الداخلي
- يوجد على الغشاء الخارجي للمايتوكندريا والبلاستيدات بعض البروتينات الناقلة porins ، مماثلة لتلك الموجودة على الغشاء الخلوي للبكتيريا.[2][3][4]
- تم العثور على الغشاء الدهني كارديوليبين الموجود فقط في الغشاء الداخلي للمايتوكندريا وغشاء البكتيريا.[5]
- كما تمتلكان رايبوسومات مشابهة لتلك الموجودة لدى البكتيريا.[1]
- تتشابه المايتوكندريا والبكتيريا بالعديد من الانزيمات وانظمة النقل.
- تحتوي خلايا الطحالب الخضراء على بلاستيدات تتشابه بشكل كبير مع الخلايا البكتيرية، حيث تحتوي هذه البلاستيدات على طبقة ببتيدوغلايكان بين الغشائين الخلويين فيها.
من حيث المعلومات والمادة الوراثية
- تمتلك كل من هذه العضيات مركب حمض نووي ريبوزي منقوص الأكسجين دائري منفرد، يختلف عن ذلك الموجود في نواة الخلية، تماما مثل الDNA الموجود في الخلية البكتيرية شكلا وتركيبا.[1]
- كما يتشابهون ايضا بتسلسل ال حمض نووي ريبوزي ريبوسومي، الحساسية لمثبطات ال RNA، وحجم الرايبوسومات (Ribosomes).[1]
- الجينوم (مجموعة العوامل الوراثية) بما في ذلك جينات معينة، تتشابه بشكل اساسي بين المايتوكندريا والبكتيريا الريكتسية.[6]
- مقارنات الجينوم(مجموعة العوامل الوراثية) تشير إلى أن البكتيريا الزرقاء ساهمت في الأصل الجيني للبلاستيدات.[7]
- تحليل تسلسل الحمض النووي والتقديرات تشير إلى أن النشوء والتطور DNA يحتوي على الجينات التي ربما جاءت من البلاستيدات .
- بعض البروتينات المشفرة في النواة تنتقل إلى العضيات، كما ان الجينوم الخاص بكلا المايتوكندريا والبلاستيدات يكون صغيرمقارنة بجينوم البكتيريا. وهذا يتفق مع الاعتماد المتزايد على الخلية المضيفة حقيقية النواة بعد تشكيل تعايش داخلي. فيتم فقد معظم الجينات في جينوم هذه العضية أو نقلها إلى النواة. وتقع معظم الجينات اللازمة لوظيفة الميتوكوندريا في النواة. كل ذلك ينشأ من التعايش الداخلي للبكتيريا في الخلية المضيفة حقيقية النواة.
من حيث الانقسام
- اثناء انقسام الخلية تنقسم عضياتها وتتضاعف بالمثل من خلال اوامر الخلية، لكن تجديد وتكاثر المايتوكندرياوالبلاستيدات اثناء الانقسام الخلوي يتم من خلال عملية شبيهة بالانشطار الثنائي الخاص بالخلية البكتيرية.
- إذا تم اخراج المايتوكندريا أو البلاستيدات من الخلية، فليس لديهم الوسيلة لاعادة التكاثر.[8] على سبيل المثال: في بعض الطحالب، مثل الحنديرة، يمكن تدميرالبلاستيدات من خلال استخدام بعض المواد الكيميائية أو حرمانها من الضوء لفترة طويلة دون أن يؤثر ذلك على الخلية. في مثل هذه الحالة، فإن البلاستيدات لا تتجدد.
تاريخ النشوء التعايشي
نظرية النشوء التعايشي: تم وضع النظرية على يد عالم النبات الروسي كونستانتين عام 1910، على الرغم من وضع اساسياتها قبل خمس سنوات، وكان عمل كونستانين مشابها لعمل عالم النبات اندرياس شمبر، الذي قد لاحظ ان تقسيم البلاستيدات الخضراء تشبه إلى حد كبير تقسيم نوع من انواع البكتيريا الخضراء المزرقة عام 1884وكان قد اقترح مبدايا أن النباتات الخضراء قد نشأت من اتحاد تكافلي لاثنين من الكائنات الحية. مدد ايفان والين فكرة النشوء التعايشي إلى المايتوكندريا لاحقا في العشرينيات من القرن الماضي، بعد دلك قام عالم النبات الروسي بوريس كوزو بولينيسكي بشرح النظرية عن طريق التطور الدارويني في كتابه الدي الفه عالم 1924 (النشوء التعايشي )، ولكن كل النظريات لم يلق لها بال وتم تجاهلها. اظهرت الميكروسكوبات تفاصيا أكثر لكل من البكتيريا الزرقاء والبلاستيدات الخضراء (مثل ابحاث هانز ريس التي نشرت في عام 1961، جنبا إلى جنب مع اكتشاف أن البلاستيدات والميتوكوندريا تحتوي على حمضها النووي الخاص بداخلها (الذي تم التعارف على انها المادة الوراثية في الكائن الحي ) والدي ادى إلى قيام الفكرة في ستينيات القرن الماضي حيث ان لكليهما القدرة على الانقسام المتساوي. رُفضت الفرضية التي قدمتها باحثة البيولوجية التطوّرية الأميركية لين مارغليوس من قرابة خمس عشرة دوريّة علميّة عام 1966، التي تقول بأنّ الجينوم في عضيات الخلية كالميتوكندريا والبلاستيدات الخضراء هو بقايا كائنات قديمة وحيدة الخلية كالبكتيريا، والتي اتحدت على مرّ العملية التطوّرية لتشكّل كائنات عديدة الخلايا ومعقدة. سبب رفض هذه الفرضية هي شيوع فكرة أنّ الميتوكندريا والبلاستيدات الخضراء لا تحتوي عل الحمض النووي الخاص بها في دلك الوقت، وعليه كانت هذه النظرية حول أصل حقيقيات النوى ثوريّة في ذلك الوقت إلى حدٍ كبيرٍ. قاومت لين مرغيليوث رفض الدوريّات العلمية لفرضيتها بعناد واستطاعت إقناع دوريّة (صحيفة الاحياء النظرية ) وتم نشرها بعد دلك بعام واحد. ويُعتقد أنه عبر آلآف السنين ادت هذه المعايشات الداخلية بنقل بعض من الحمض النووي الخاص بها إلى نواة الخلية المضيفة وذلك خلال الانتقال التطوري من مجموعة متكافلة إلى خلية حقيقية النواة.
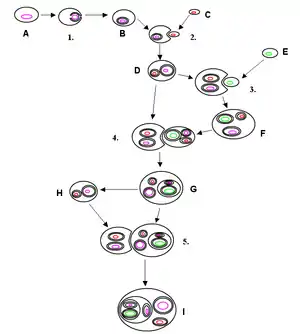
النشوء التعايشي الثانوي
يقوم النشوء التعايشي الأساسي على الإحاطة بنوع من البكتيريا، وذلك بواسطة كائن حي حر. يحدث النشوء التعايشي الثانوي عندما يتم اجتياح منتج النشوء التعايشي الأساسي بواسطة كائن آخر حر وذي نواة حقيقة. وقد حدث هذا التعايش الثانوي عدة مرات، مما أدى إلى ظهور مجموعات مختلفة للغاية من الطحالب وغيرها من حقيقيات النواة. وتستطيع بعض الكائنات الحية الاستفادة من مثل هذه العملية، وذلك من خلال اجتياح الطحالب واستخدام منتجاتها من التمثيل الضوئي، ولكن عند موت الطحلب الذي تم اجتياحه أو فقده يعود المضيف إلى حالته الحية. أصبح النشوء التعايشي أمرًا ضروريًا ولازمًا للعضيات، حيث لا يمكنها البقاء على قيد الحياة بدون وجوده. وقد لاحظ أوكاموتواوإينوي عام (2005) إمكانية تعايش داحلي واحد في هذه العملية.[9] ويتصرف وحيد الخلية الذي يتغذى على غيره هاتينا مثل الحيوان المفترس حتى يبتلع الطحالب الخضراء، والتي تفقد سياطها وهيكلها الخلوي بسبب ذلك، في حين يتحول هاتينا، والذي أصبح مضيفًا الآن، إلى التغذية الضوئية ويفقد جهاز التغذية الخاص به. وتترك عملية النشوء التعايشي أثرها التطوري داخل التضاريس الفريدة لأغشية البلاستيد. وتحاط البلاستيدات الثانوية بثلاث ( طحالب يوجلينية وبعض الطحالب الدوارة ) أو أربعة أغشية (هابتوفيت، هيتركونا، كريبتوفيت وكلوراشنيوفيت ). ويعتقد أن الغشائين الإضافيين يعملان على التوافق مع غشاء البلازمة الخاص بالطحلب المجتاح ومع غشاء الخلية المضيفة. ويتمثل الحصول على النشوء التعايشي لخلية حقيقية النواة في الكريبتوفيتس، حيث توجد نواة من الطحالب الحمراء المتكافلة نكليومورف بين غشائين بلاستيديين داخليين وغشائين خارجيين وعلى الرغم من تنوع الكائنات التي تحتوي على البلاستيدات، تشير المورفولوجيا ، الكيمياء الحيوية، تنظيم الجينوم والنسالة الجزيئية من الحمض الريبي النووي RNAs و البروتينات إلى أصل واحد من كل البلاستيدات الموجودة.[10][11] ولكن لاتزال هذه النظرية محل جدال ونقاش وتمتلك بعض الأنواع بما في ذلك القمل البشري كروموسومات متعددة في الحبيبات الخيطية. ويشير هذاإلى جانب تطورات سلالات الجينات المشفرة داخل الحبيبات الخيطية إلى أنه قد تم الحصول على جذور الميتوكوندريا عدة مرات قبل ذلك وليس مرة واحدة.[12]
نقاط ضعف النظرية
هنالك عدد من النقاط التي تصعف أو تبطل نظرية النشوء التعايشي:
1- على الرغم من وجود بعض الخصائص المشتركة بين أغشية البكتيريا والمايتوكندريا، إلا أنه يوجد أيضا اختلافات لعل أبرزها هو قدرة المضادات الحيوية على التمييز بين أغشية البكتيريا والمايتوكندريا، وهذا يدعم الفكرة القائلة ( أن الأغشية الخلوية يختلف تركبيها من غشاء إلى أخر ويمكن تحديدها من تحديدها من قبل وكلاء مثل المضادات الحيوية)، ومن الصعب محاولة تغيير المكونات الأساسية لأغشية البكتيريا، والعثور على نظير مماثل لأغشية الميتوكوندريا دون أي تغيير على سلامة الغشاء. هذا الاختلاف في بنية الغشاء أحد نقاط التي تخالف نظرية النشوء التعايشي.[13]
2- المايتوكنددريا والبكتيريا لديها رايبوسومات مصنوعة من بروتين و (RNA)؛ لتحفيز إنتاج البروتينات، وكان أول من اقترح نظرية النشوء التعايشي، افترض أن الرايبوسومات وجدت في شكلين فقط، الأول: رايبوسومات ذات ذات أشكال متنوعة صغيرة 70S موجودة في الخلايا بدائية النوى، و الثاني: رايبوسومات أكبر 80S موجودة في العصارة الخلوية في الخلايا حقيقية النوى.ووفقا لنظرية النشوء التعايشي كان من المفروض أن الرايبوسومات الموجودة في مايتوكندريا الثدييات أن تتطايق رايبوسومات الخلايا بدائية النوى 70S، لكن لكن تركيب رايبوسومات مايتوكندريا الثدييات، ال(RNA)وتسلسل الحموض الأمينية مخالف تماما لتركيب رايبوسومات الخلايا بدائية النوى.[14]
3-تسلسل جينوم المايتوكندريا يمكن أن يحتوي على خليط من الخلايا حقيقية النوى، الخاليا بدائية النوى والخلايا القديمة.وهذا يختلف مع ما هو متوقع إذا افترضنا أن أصل المايتوكوندريا فقط خلايا بدائية النوى. [15]
4- إذا افترضنا أن البلاستيدات في يوم ما كانت مستقلة، فبالتالي يمكن أن يكون هنالك ناتج واحد فقط إذا ابتلعتها الخلية الأم، وهو أن يتم هضمها من قبل الخلية الأم واستخدامها كغذاء، وهاذا ما يجب أن يحدث لأنه لو افترضنا في هذه الحالة أن الخلية الأم بالخطأ أدخلت مثل هذه الخلية من الخارج بدلا من التهامها عمدا، مع ذلك فإن الإنزيمات الهاضمة في الخلية الأم قد دمرتها، وهاذا تناقض واضح مع إدعاء بعض أنصار التطور " الإنزيمات الهاضمة قد دمرت" لأنه لو اختفت الإنزيمات الهاضمة في الخلية الأم، لأصبحت هذه الخلية ميتة بسبب نقص التغذية. [16]
5- يتم ترميز جميع المخططات لجميع العضيات داخل الخلية في الحمض النووي، وإذا كانت الخلية الأم ستستخدم الخلية التي ابتلعتها كعضيات، فإنه سيكون من الضروري أن تتواجد المعلومات المتعلقة بترميزها في (DNA) الخلية الأم، وال(DNA) للخلايا التي تم ابتلاعها يجب أن تمتلك المعلومات التي تنتمي للخلية الأم، وليس هذا فقط المستحيل، بل يجب أن يوجد جزيئان متممان من ال (DNA) ينتميان للخلية الأم، والخلية التي ابتلعت أيضا يجب أن تكون متوافقة مع بعضها البعض بعد ذلك وهو مستحيل حدوثه بوضوح.[16]
6- مسألة كيف بدأت الخلايا النباتية بعملية البناء الضوئي هل من الممكن لمثل عملية معقدة كالبناء الضوئي أن تكون ناتج لعمليات طبيعية كالتي تقوم عليها نظرية التطور؟ وفقا لسيناريو التطور، من أجل القيام بعملية البناء الضوئي الخلايا النباتية تبتلع الخلايا البكتيرية التي مكنتها من القيام بعملية البناء الضوئي، وحولتها إلى بلاستيدات، كيف تعلمت البكتيريا تنفيذ عملية معقدة كالبناء الضوئي؟ لماذا لم تبدأ هذه العملية من قبل؟ [17]
من المعايش الجواني إلى العضيات الخلوية
طبقا لكييلنغ وأرشيبولد، فإن الطريقة الشائعة للتفريق بين العضيات الخلوية وبين المعايشات الجوانية هو حجم الجينوم الأقل. مع تطور المعايش الجواني إلى عضية خلوية، تنتقل معظم جيناته إلى جينوم الخلية المضيفة (المجموع الوراثي). تحتاج الخلية المضيفة والعضية الخلوية إلى تطوير آلية نقل تسمح بعودة نواتج البروتين التي تحتاجها العضيات الخلوية ولكنها تُصنع الآن في الخلية.[18] البكتريا الزرقاء ومتقلبات ألفا هي الكائنات الحرة الأكثر قرابة للبلاستيدات والميتوكوندريا على التوالي. يحافظ كل من البكتريا الزرقاء ومتقلبات ألفا على جينوم كبير الحجم يسمح بتشفير آلاف البروتينات. تتميز البلاستيدات والميتوكوندريا بانخفاض هائل في حجم الجينوم عند مقارنتها بمثيلاتيها في البكتريا.[19] يتراوح حجم جينوم الكلوروبلاست في الكائنات الحية التي تقوم بالبناء الضوئي بين 120 ألف و200 ألف مما يسمح بتشفير 20-200 بروتين، بينما يبلغ حجم جينوم الميتوكوندريا في البشر حوالي 16 ألف ليسمح بتشفير 37 جينا، منهم 13 بروتين. تُظهر المقارنة مع البكتريا الزرقاء الأكثر قرابة لها من جنس المتعاقبة الحبيبية (والتي تملك جينوما يبلغ حجمه 3 ملايين و3300 جينا) أن الكروماتوفور (الخلايا الصبغية المستجيبة للضوء) شهدت انكماشا هائلا في حجم الجينوم. احتوت الخلايا الصبغية المستجيبة للضوء على جينات مسؤولة عن التركيب الضوئي ولكنها كانت غير فعالة في جينات قادرة على القيام بوظائف تركيب ضوئي أخرى. يشير هذا الاكتشاف إلى أن خلايا المعايش الجواني تعتمد بصورة كبيرة على الخلايا المضيفة من أجل بقائها وآليات نموها. بالتالي، فإنه تم الكشف عن أن هذه الخلايا الصبغية المستجيبة للضوء غير فعالة للأغراض الخاصة بالعضيات الخلوية خاصة عند مقارنتها بالميتوكوندريا أو البلاستيدات. قد يكون هذا الفصل هو ما حفز عملية التطور المبكرة للعضيات الخلوية المسؤولة عن التركيب الضوئي. حدث فقد العديد من جينات المعايش الجواني في نقطة مبكرة جدا في الزمن التطوري.[20] عند أخذ جينوم المعايش الجواني الكامل في الاعتبار، فإنه هناك ثلاثة مصائر محتملة للجينات بمرور الزمن التطوري. المصير الأول هو فقد الجينات الفعالة الإضافية، إذ تفقد الخلية الجينات الممثلة بالفعل في نواة الخلية. يتضمن المصير الثاني نقل الجينات الأفقي إلى نواة الخلية.[21] يمكن إرجاع فقد استقلالية المعايشات الجوانية مع الخلية المضيفة بصورة أساسية إلى نقل الجينات إلى النواة. وبما أن جينوم العضيات الخلوية قد شهد انخفاضا كبيرا مع مرور الزمن التطوري، فإن الجينات النووية قد تمددت وأصبحت أكثر تعقيدا. كنتيجة لذلك، فإن العديد من عمليات الميتوكوندريا والبلاستيدات تقوم بها نواتج من تشفير الجينات والتي تحدث في النواة. بالإضافة إلى ذلك، فإن العديد من الجينات النووية الناشئة من المعايشات الجوانية قد اكتسبت وظائف غير مرتبطة بالعضيات الخلوية. آليات انتقال الجينات ليست مفهومة تماما، إلا أنه هناك العديد من الفرضيات التي تحاول تفسير هذه الظاهرة. تتضمن فرضية الدنا المتمم استخدام الحمض النووري الريبوزي الرسول mRNA لنقل الجينات من العضيات الخلوية إلى النواة حيث يتم تحويلها إلى حمض نووي ريبوزي منقوص الأكسجين متمم cDNA ويتم تضمينها في الجينوم. تعتمد فرضية الدنا المتمم على دراسات الجينوم في النباتات المزهرة.[22] منذ أن تم الإعلان عن فرضية الدنا المتمم، روجعت الفرضية إذ أليس من المحتمل أن يقوم الدنا المتمم في المتيوكوندريا بالاتحاد مع الجينوم الميتوكوندري، كما أن مواقع الإقران الميتوكوندرية لن تكون موجودة في الجينوم الميتوكوندري. بالتالي فإن أي عملية نقل جيني إلى النواة ستؤدي إلى انعدام مواقع الإقران على الميتوكوندريا. فرضية الكتلة السائلة هي فرضية بديلة لفرضية الدنا المتمم والتي تنص على أن الحمض النووري الريبوزي منقوص الأكسجين الهارب –وليس الدنا الحمض النووي الريبوزي الرسول- هو المسؤول عن آلية نقل الجينات إلى النواة.[23]
قراءة إضافية
Margulis, L.,and D. Sagan. Acquiring Genomes: A Theory of the Origins of Species. New York: Basic Books,2003.
Poole, A.M., and D. Penny. Evaluating hypotheses for the origin of eukaryotes. BioEssays 29 (2007): 500.
مراجع
- Becker's World of the Cell, Book, Edition (8th Edition) 8th Edition by Jeff Hardin (Author), Gregory Paul Bertoni (Author), Lewis J. Kleinsmith (Author) page 89
- Fischer K, Weber A, Brink S, Arbinger B, Schünemann D, Borchert S, Heldt HW, Popp B, Benz R, Link TA (1994). "Porins from plants. Molecular cloning and functional characterization of two new members of the porin family". J Biol Chem 269 (41): 25754–25760. PMID 7523392
- Zeth K, Thein M (2010). "Porins in prokaryotes and eukaryotes: common themes and variations". Biochem J 431 (1): 13–22. doi:10.1042/BJ20100371. PMID 20836765
- Fairman JW, Noinaj N, Buchanan SK (2011). "The structural biology of β-barrel membrane proteins: a summary of recent reports". Current Opinion in Structural Biology21 (4): 523–531. doi:10.1016/j.sbi.2011.05.005. PMC 3164749. PMID 21719274
- Mileykovskaya E, Dowhan W (2009). "Cardiolipin membrane domains in prokaryotes and eukaryotes". Biochim Biophys Acta 1788 (10): 2084–2091.doi:10.1016/j.bbamem.2009.04.003. PMC 2757463. PMID 19371718
- Andersson SG, Zomorodipour A, Andersson JO, Sicheritz-Pontén T, Alsmark UC, Podowski RM, Näslund AK, Eriksson AS, Winkler HH, Kurland CG (1998). "The genome sequence of Rickettsia prowazekii and the origin of mitochondria". Nature 396 (6707): 133–140. doi:10.1038/24094. PMID 9823893
- Dagan T, Roettger M, Stucken K, Landan G, Koch R, Major P, Gould SB, Goremykin VV, Rippka R, Tandeau de Marsac N, Gugger M, Lockhart PJ, Allen JF, Brune I, Maus I, Pühler A, Martin WF (2013). "Genomes of Stigonematalean cyanobacteria (subsection V) and the evolution of oxygenic photosynthesis from prokaryotes to plastids". Genome Biol Evol 5 (1): 31–44. doi:10.1093/gbe/evs117. PMC 3595030. PMID 23221676
- Wise, Robert R; Hoober,, J. Kenneth (2007). Structure and function of plastids. Berlin: Springer. p. 104. ISBN 9781402065705.
- Okamoto, Noriko; Inouye, Isao (2005). "A Secondary Symbiosis inProgress?" (PDF).Science 310 (5746):287. doi:10.1126/science.1116125. PMID 16224014. Retrieved15 February 2015.
- McFadden GI, van Dooren GG (July 2004). "Evolution: red algal genome affirms a common origin of all plastids". Curr. Biol. 14 (13):R5146.doi:10.1016/j.cub.2004.06.041. PMID 15242632
- Gould SB, Waller RF, McFadden GI (2008). "Plastid evolution". Annu Rev Plant Biol59 (1): 491–517. doi:10.1146/annurev.arplant.59.032607.092915. PMID 18315522
- Georgiades, K. and Raoult, D. (2011). "The rhizome of Reclinomonas americana, Homo sapiens, Pediculus humanus and Saccharomyces cerevisiae mitochondria". Biology Direct6: 55. doi:10.1186/1745-6150-6-55
- Dr. Ferguson(2012) Criticism. https://theendosymbiotichypothesis.wordpress.com/controversy-2/ نسخة محفوظة 2021-01-10 على موقع واي باك مشين.
- Daniel C. Criswell(2009) A Review of Mitoribosome Structure and Function does not Support the Serial Endosymbiotic Theory. www.answersingenesis.org/contents/379/arj/v2/Serial_Endosymbiotic_Theory.pdf
- Backgrounder: Some challenges offered for Lynn Margulis’s endosymbiosis theory | Uncommon Descent نسخة محفوظة 10 سبتمبر 2016 على موقع واي باك مشين.
- Wallace-Sanders-Ferl, Biology: The Science of Life, 4th edition, Harper Collins College Publishers, p. 94
- Milani, Bradshaw, Biological Science, A Molecular Approach, D. C.Heath and Company, Toronto, p. 158
- Michael Syvanen, Clarence I. Kado Horizontal Gene Transfer Academic Press, p. 405 (ردمك 978-0126801262)
- Keeling, P. J.; Archibald, J.M. (2008). "Organelle evolution: what's in a name?". Current Biology. 18 (8): 345–347. doi:10.1016/j.cub.2008.02.065. PMID 18430636. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lila Koumandou, V.; Nisbet, R. Ellen R.; Barbrook, Adrian C.; Howe, Christopher J. (2004). "Dinoflagellate chloroplasts – where have all the genes gone?". Trends in Genetics. 20 (5): 261–267. doi:10.1016/j.tig.2004.03.008. PMID 15109781. مؤرشف من الأصل في 14 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Taanman, Jan-Willem (1999-02-09). "The mitochondrial genome: structure, transcription, translation and replication". Biochimica et Biophysica Acta (BBA) - Bioenergetics. 1410 (2): 103–123. doi:10.1016/S0005-2728(98)00161-3. مؤرشف من الأصل في 24 أبريل 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Nowack, E.C.; Melkonian, M.; Glockner, G. (2008). "Chromatophore genome sequence of Paulinella sheds light on acquisition of photosynthesis by eukaryotes". Current Biology. 18 (6): 410–418. doi:10.1016/j.cub.2008.02.051. PMID 18356055. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Barbrook, Adrian C.; Howe, Christopher J.; Purton, Saul (2006). "Why are plastid genomes retained in non-photosynthetic organisms?". Trends in Plant Science. 11 (2): 101–108. doi:10.1016/j.tplants.2005.12.004. PMID 16406301. مؤرشف من الأصل في 14 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء التطوري
بوابة علم الأحياء التطوري
