مستقبل غابا-أ
مستقبل حمض الغاما-أمينوبيوتيريك هو مستقبل أيونوتروبي وقناة أيونية. تستهدف هذه المستقبلات حمض الغاما-أمينوبيوتيريك، الذي يعتبر الناقل العصبي المثبط الرئيسي في الجهاز العصبي المركزي. وعند تنشيط تلك المستقبلات فإنها تضخ الكلوريد عن طريق قنواتها الأيونية، مما يؤدي إلى فرط الاستقطاب للأعصاب. ويتسبب هذا في تثبيط انتقال النواقل العصبية بتقليل احتمالية حدوث جهد الفعل. يختلف جهد الاعتكاسفي مستقبلات GABAA عن نظيراتها في GABAB والذي يساوي في المحلول العادي -70 ميللي فولت.
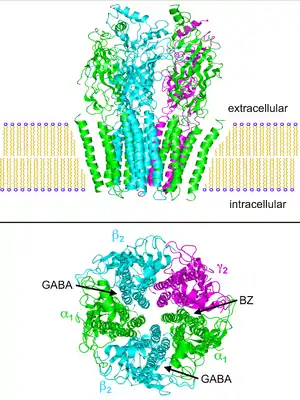
على اليمين: خمس وحدات فرعية رتبت بشكل متناظر حول مسام توصيل أنيون الكلوريد المركزي . لا يتم تصوير الحلقات خارج الخلية من أجل الوضوح.
يعتبر الموقع النشط لمستقبلات GABAA حمض الغاما-أمينوبيوتيريك موقع ارتباط للعديد من الأدوية مثل موسكيمول وجابوكسادول وبيكوسوللين.يحتوي البروتين أيضًا على مواقع تفارغية والتي تعدل من نشاط تلك المستقبلات بصورة غير مباشرة. كما تعتبر تلك المواقع التفارغية وجهة ارتباط للعديد من الأدوية مثل البنزوديازيبين وغير البنزوديازيبينات والاسترويدات العصبية والباربتيورات والإيثانول[4] ومواد التخدير المُستنشقة والبيكروتوكسين .[5]
توجد مستقبلات غابا في جميع الكائنات الحية التي لديها جهاز عصبي. كما يمكن العثور عليها إلى حد محدود في الأنسجة غير العصبية. ونظرًا لتوزيعها على نطاق واسع داخل الجهاز العصبي في الثدييات فإنها تلعب دورًا في جميع وظائف الدماغ تقريبًا.
هدف البنزوديازيبين
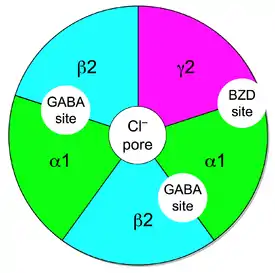
يعتبر المركب الأيونوتروبي لمستقبلات GABA وجهة لارتباط البنزوديازيبين. إلا أن البنزوديازيبينات لا ترتبط بنفس مواقع ارتباط حمض الغاما-أمينوبيوتيريك والتي ترتبط بين موقعي α و β ولكنها ترتبط بموقع خاص بها على السطح الخارجي لوحدتي α وγ.[6][7] بينما نجد أن معظم مستقبلات GABA عالية الحساسية للبنزوديازيبينات (خاصةً تلك المحتوية على وحدات α1-, α2-, α3-, or α5) إلا أنه توجد بعض الوحدات الغير حساسة على الإطلاق لعائلة البنزوديازيبينات (خاصةً تلك المحتوية على وحدات α4- or α6)، [8] ولكنها حساسة للمواد الأخرى التي تستهدف تلك المستقبلات مثل الاسترويدات العصبية و الباربتيورات والإيثانول وتباعًا لهذا، أوصى الاتحاد الدولي للصيدلة الأساسية والطبية بتغيير تسمية مصطلح "مستقبلات البنزوديابين" GABA/BZ ومستقبلات أوميغا إلى مصطلح مواقع ارتباط البنزوديازيبين.[9]
ولكي تكون مستقبلات GABA حساسة لعمل البنزوديازيبينات فإنها تحتاج إلى أن تحتوي على وحدتي α و γ حيث ترتبط البنزوديازبينات. وبمجرد حدوث الارتباط، تقوم البنزوديازيبينات بغلق المستقبلات بشكل كلي للوقاية من التقارب العالي لمستقبلات غابا للناقل العصبي حمض الغاما-أمينوبيوتيريك، وزيادة وتيرة انفتاح قناة أيون الكلوريد المرتبطة مما يؤدي إلى فرط الاستقطاب. يعزز هذا من التأثير المثبط لحمض الغاما-أمينوبيوتيريك مما يؤدي إلى تأثيرات مهدئة ومضادة للقلق.
تختلف البنزوديازيبينات بين بعضها البعض في درجة التقارب لمستقبلات GABA. وهذا يعني أن خصائصها الدوائية تختلف باختلاف النوع الفرعي. على سبيل المثال، ترتبط البنزوديازيبينات المرتبطة بالمستقبلات المحتوية على الوحدات الفرعية α1 و/أو α5 بتأثيرات مثل التهدئة والرنح وفقدان الذاكرة، بينما ترتبط البنزوديازيبينات المرتبطة بالمستقبلات المحتوية على الوحدات الفرعية α2 و/أو α3 بتأثير مضاد للقلق.[10] يحدث التأثير المضاد للصرع عند تحفيز أي من مستقبلات GABA بوحداتها الفرعية المختلفة، لكن الأبحاث الحالية في هذا المجال تركز على إنتاج منبهات α2 الانتقائية مثل مضادات الاختلاج التي تفتقر إلى الآثار الجانبية للأدوية القديمة مثل التخدير وفقدان الذاكرة.
تختلف مواقع ارتباط البنزوديازيبينات عن تلك الخاصة بالباربتيورات، كما يؤدي كل منهم إلى تأثير مختلف عن الآخر عند حدوث الارتباط على موقع GABAA الخاص بمستقبل GABA، إذ تزيد البنزوديتزيبينات من وتيرة انفتاح قنوات الكلوريد،[11]
بينما تزيد الباربيتورات من مدة انفتاح تلك القنوات.[12]
وبما أن هذه الآثار التماثلية منفصلة، فبإمكانها أن تحدث في نفس الوقت، وبالتالي فإن الجمع بين البنزوديازيبينات والباربيتورات يؤدي إلى تأثير تآزري قوي، والذي قد يكون خطيرًا إذا لم يتم التحكم في الجرعات بشكل صارم.
تجدر الإشارة إلى أن بعض منبهات GABAA مثل موسكيمول وجابوكسادول وبيكوسوللين بإمكانها أن ترتبط إلى نفس مواقع ارتباط حمض الغاما-أمينوبيوتيريك، مسببةً آثار مقاربة-لكنها لا تطابق- تلك الناتجة عن البنزوديازيبينات.
التركيب والوظيفة
استند الفهم الهيكلي لمستقبلات GABA في البداية إلى نماذج التماثل التي تم الحصول عليها باستخدام هياكل الكريستال للبروتينات المتجانسة مثل بروتينات أستيل كولين ومستقبلات النيكوتينيك أستيل كولين كنماذج.[14][15] تم التوصل أخيرًا إلى بنية مستقبلات GABA مع الكسف عن التركيب الكريستالي ذي التجانس الخماسي لوحدة β3 الخاصة بمستقبل GABAA .[16][17][18][19][20]
الوحدات الفرعية
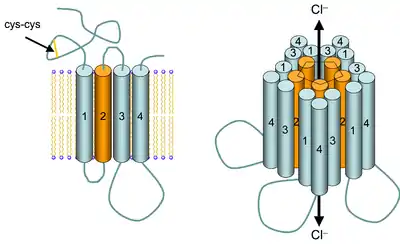
مستقبلات غابا هي أعضاء في عائلة "CYS-LOOP" الكبيرة المرتبطة بشكل كبير بالمستقبلات الأيونوتروبية والتي تشمل أيضا مستقبلات الأسيتيل كولين، مستقبلات الجلايسين، ومستقبلات 5HT3. توجد هناك العديد من الأشكال الإسوية للوحدات الفرعية لمستقبل GABAA، والتي تحدد تقارب المستقبلات، وانفتاح القنوات، والتوصيل وغيرها من الخصائص.[21]
توجد الوحدات في البشر كالآتي:[22]
- ست وحدات من النوعα subunits (GABRA1، GABRA2، GABRA3، GABRA4، GABRA5، GABRA6)
- ثلاث وحدات β (GABRB1 GABRB2، GABRB3
- ثلاث وحدات γ (GABRG1 GABRG2، GABRG3
- بالإضافة إلى δ (GABRD), an ε (GABRE)و a π (GABRP) و a θ (GABRQ)
كما توجد هناك ثلاث مستقبلات تحتوي على وحدات فرعيةρ GABRR1، GABRR2، GABRR3؛ ومع ذلك لا يتم ذكر مثل تلك الوحدات مع مستقبلات GABA المذكورة بالأعلى، [23] لكنها تتجانس مكونةً مستقبلات GABAA-ρ ( كانت تُسمى فيما مضى مستقبلات GABAC، لكن أهملت هذه التسمية حاليًا[24]).
يمكن أن تتجدد خمس وحدات فرعية بطرق شتى لتكوين قناة GABAA. إذ أن الحد الأدنى لتكوين قناة GABA أيونية أن تحتوي على وحدتي α و β.[25] لكن النوع الشائع من المستقبلات الموجودة في المخ هو تلك التي تحتوي على وحدتي α ووحدتي β ووحدة γ (α1β2γ2).[22] يمكن للمستقبل أن يرتبط بجزيئين من حمض الغاما-أمينوبيوتيريك، [26] بين وحدتي α و β.[22]
الربيطات
عثر على العديد من الربيطات التي ترتبط بمستقبلات GABA بخلاف حمض الغاما-أمينوبيوتيريك وتحدث تأثير مستقل بها. يمكن للربيطة أن تمتلك خاصية أو أكثر من تلك الأنواع والتي لسوء الحظ دائمًا ما تخطيء المؤلفات في التمييز بين تلك الأنواع.
الأنواع
- الناهضة والمناهضة: ترتبط بالموقع الرئيسي للمستقبلات (الموقع الذي يربط به حمض الغاما-أمينوبيوتيريك عادةً، وُيشار إليهِ أيضا باسم "الموقع النشط" أو " الموقع الناهض"). تنشط الناهضات المستقبلات، مما يؤدي إلى زيادة توصيل الكلوريد. وعلى الرغم من عدم وجود تأثير مستقل للمناهضات، إلا أنها تتنافس مع حمض الغاما-أمينوبيوتيريك للارتباط بالمستقبلات وبالتالي تمنع عملها، مما يؤدي إلى انخفاض توصيل الكلوريد.
- الناهضات غير المباشرة: ترتبط تلك الناهضات بالمواقع الفرعية على المستقبلات وتفعل المستقبلات في غياب الارتباط بالموقع الرئيسي "النشط".
- المعدلات غير المباشرة الأولية: ترتبط تلك الناهضات بالمواقع الفرعية على المستقبلات محدثةً تأثير إيجابي أو سلبي أو تعادلي، مما يؤدي إلى زيادة كفاءة الموقع الرئيسي للارتباط فيترتب على ذلك ارتفاع أو انخفاض توصيل الكلوريد. لا تؤثر الناهضات غير المباشرة الثانوية على التوثيل لكنها تشغل مواقع الارتباط فقظ.
- المعدلات غير المباشرة الثانوية: ترتبط تلك الناهضات بالموقع الفرعي على المستقبلات وتعدل من تاثير الناهضات غير المباشرة الأولية.
- مضادات انفتاح القنوات: تُطيل من فترة ارتباط الربيطة بالمستقبل، وتنشيط حركية واندفاع أيونات الكلوريد في صورة تعتمد على الحساسية لتلك الأيونات.[27]
- مضادات القنوات غير التنافسية: ترتبط عند أو بالقرب من منتصف البوابة وتمنع توصيل الكلوريد من خلال القناة الأيونية.
أمثلة
- الناهضة الرئيسية: حمض الغاما-أمينوبيوتيريك وموسكيمول وجابوكسادول وبيبيريدين-4-حمض السلفونيك (ناهض جزئي)
- المناهضة الرئيسية: بيكوسوللين وجابازين.
- الناهضات غير المباشرة الموجبة: الباربتيورات والبنزوديازيبين وبعض الكربامات (مثل كاريسوبرودول وميبروبامات ولوربامات) و الثينوديازيبينات، والإيثانول (كحول)، والإتوميديت، والغلوتيثيميد، والكافالاكتونيس،
[28] والميبروبامات، والكينازولينونيس (مثل ميثاكوالون، إتاكوالون، ديبروكوالون)، والمنشطات العصبية،[29]النياسين / النيكوتيناميد،[30]البنزوديازيبينات (زولبيديم، إزوبيكلون)، والبروبوفول، والستيريبنتول، الثيانين،[31]حمض الفاليرينيك، والتخدير المستنشق، واللانثانوم.[32]
- الناهضات غير المباشرة السالبة: فلومازينيل، RO15-4513، سارمازينيل، أومنتوفلافون، والزنك.[33]
- المعدلات غير المباشرة الثانوية: إبيغالوكاتشين-3-غاليت.
- مضادات القنوات غير التنافسية: سيكتوكسين، أوينانثوتوكسين، بنتيلينتيترازول، بيكروتوكسين، ثوجون، والليندين.
التأثير
دائمًا ما تتمتع الربيطات التي تساهم في تنشيط المستقبلات بخواص مضادات القلق ومضادات الاختلاج وفقدان الذاكرة والمهدئات والمنومات والابتهاج ومرخيات العضلات. بعض الأدوية مثل الموسكيمول قد تتمتع بخواص الهلوسة. بينما تتمتع الربيطات التي تعمل على تثبيط المستقبلات بالخواص المغايرة، مثل الإصابة بالقلق والاختلاج. تجرى بعض البحوث حول التأثيرات المنشطة للذهن للمؤثرات الانتقائية غير المباشرة مثل α5IA بالإضافة إلى علاج الآثار الجانبية غير المرغوب فيها للأدوية المحفزة من نشاط حمض الغاما-أمينوبيوتيريك.[34]
أدوية جديدة
توجد هناك خاصية مفيدة للعديد من مؤثرات البنزوديازيبين غير المباشرة، وهي أنها يمكن أن تظهر خواص ارتباط انتقائية لمجموعات فرعية معينة من المستقبلات التي تضم وحدات فرعية محددة. وهذا يسمح لمؤثر واحد فقط لتحديد أيًا من مستقبلات الوحدات الفرعية كي تكون هي السائدة في مناطق معينة من الدماغ، ويقدم دليلاً على أي من مجموعات الوحدات الفرعية قد تكون مسؤولة عن الآثار السلوكية للأدوية التي تعمل في مستقبلات GABA. قد تكون لهذه الروابط الانتقائية مزايا دوائية لأنها قد تسمح بالفصل بين الآثار العلاجية المرغوبة والآثار الجانبية غير المرغوب فيها. [35] سمح باستخدام عدد قليل من الربيطات النتقائية للواحدات الفرعية في الاستخدام الطبي حتى الآن، باستثناء الزولبيديم (زولبيديم) الذي يعتبر بمثابة ربيط انتقائي ل α1. كما يتم تطوير العديد من الربيطات الانتقائية مثل عقار أديبيبلون الانتقائي لوحدة α3. توجد العديد من الأدوية التي تُجرى عليها البحوث العلمية للربيطات الانتقائية مثل:
- CL-218,872 (ناهض عالي الانتقاء لوحدة α1)
- بريتازينيل (ناهض جزئي انتقائي للأنواع الفرعية)
- اميدازينيل and L-838,417 (كلاهما ناهضان جزئيان لبعض الأنواع، إلا أنهما مناهضان ضعيفان للأنواع الأخرى)
- QH-ii-066 (ناهض كلي عالي الانتقاء للربيطات المختوية على الوحدة الفرعية α5)
- α5IA (ناهض انعكاسي انتقائي للربيطان المحتوية على الوحدة الفرعية α5)
- SL-651,498 (ناهض كلي ل α2 and α3 وناهض جزئي لα1و α5)
- 3-أسيل-4-كينولون: انتقائي ل α1 over α3 .[36]
التوزيع
كما أن مستقبلات GABA هي المسؤولة عن معظم الأنشطة الفسيولوجية لحمض الغاما-أمينوبيوتيريك في الجهاز العصبي المركزي، لذا تنتشر وحداتها الفرعية في أجزاء كثيرة من الدماغ. يمكن أن يختلف تكوين الوحدات الفرعية بصورة كبيرة بين المناطق وقد يترافق هذا مع وظائف محددة للوحدات الفرعية. في الخلايا العصبية نفسها، تختلف أنواع كثافة الوحدات الفرعية لمستقبلات GABAA بين جسم الخلية العصبية والزوائد الشجرية.[37] بالإضافة إلى هذا، توجد مستقبلات GABAA في الخلايا البينية والمشيمة وخلية الدم البيضاء والكبد والصفيحة مشاشية) والعديد من أنسجة جهاز الغدد الصماء. تختلف انواع الوحدات الفرعية في أنسجة السرطان، كما وُجد أن بإمكان مستقبلات GABAA التأثير على تكاثر الخلايا.[38]
انظر أيضًا
مراجع
- "Diazepam-bound GABAA receptor models identify new benzodiazepine binding-site ligands". Nat. Chem. Biol. 8 (5): 455–464. March 2012. doi:10.1038/nchembio.917. PMC 3368153. PMID 22446838. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Molecular modelling of the GABAA ion channel protein". J. Mol. Graph. Model. 25 (5): 721–30. January 2007. doi:10.1016/j.jmgm.2006.06.001. PMID 16877018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Structural Determinants for High-Affinity Zolpidem Binding to GABA-A receptors". Mol. Pharmacol. 71 (1): 38–46. January 2007. doi:10.1124/mol.106.029595. PMC 2583146. PMID 17012619. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Ethanol acts directly on extrasynaptic subtypes of GABAA receptors to increase tonic inhibition". Alcohol. 41 (3): 211–21. May 2007. doi:10.1016/j.alcohol.2007.04.011. PMC 2040048. PMID 17591544. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Johnston GAR (1996). "GABAA Receptor Pharmacology". Pharmacology and Therapeutics. 69 (3): 173–198. doi:10.1016/0163-7258(95)02043-8. PMID 8783370. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Sigel E (August 2002). "Mapping of the benzodiazepine recognition site on GABAA receptors". Curr Top Med Chem. 2 (8): 833–9. doi:10.2174/1568026023393444. PMID 12171574. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Akabas MH (2004). "GABAA receptor structure-function studies: a reexamination in light of new acetylcholine receptor structures". Int. Rev. Neurobiol. 62: 1–43. doi:10.1016/S0074-7742(04)62001-0. ISBN 978-0-12-366862-2. PMID 15530567. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Identification of a residue in the gamma-aminobutyric acid type A receptor alpha subunit that differentially affects diazepam-sensitive and -insensitive benzodiazepine site binding". J. Neurochem. 88 (6): 1431–8. March 2004. doi:10.1046/j.1471-4159.2003.02264.x. PMID 15009644. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "International Union of Pharmacology. XV. Subtypes of gamma-aminobutyric acidA receptors: classification on the basis of subunit structure and receptor function". Pharmacol. Rev. 50 (2): 291–313. 1 June 1998. PMID 9647870. مؤرشف من الأصل في 14 مارس 2009. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Atack, JR (2003). "Anxioselective compounds acting at the GABA(A) receptor benzodiazepine binding site". Current drug targets. CNS and neurological disorders. 2 (4): 213–32. doi:10.2174/1568007033482841. PMID 12871032. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Structural mechanisms underlying benzodiazepine modulation of the GABAA receptor". J. Neurosci. 28 (13): 3490–9. March 2008. doi:10.1523/JNEUROSCI.5727-07.2008. PMC 2410040. PMID 18367615. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Differential regulation of gamma-aminobutyric acid receptor channels by diazepam and phenobarbital". Ann. Neurol. 25 (3): 213–20. March 1989. doi:10.1002/ana.410250302. PMID 2471436. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Diazepam-bound GABAA receptor models identify new benzodiazepine binding-site ligands". Nat. Chem. Biol. 8 (5): 455–464. March 2012. doi:10.1038/nchembio.917. PMC 3368153. PMID 22446838. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Comparative models of GABAA receptor extracellular and transmembrane domains: important insights in pharmacology and function". Mol Pharmacol. 68 (5): 1291–300. November 2005. doi:10.1124/mol.105.015982. PMID 16103045. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Modeling the closed and open state conformations of the GABA(A) ion channel-plausible structural insights for channel gating". J Chem Inf Model. 52 (11): 2958–69. November 2012. doi:10.1021/ci300189a. PMID 23116339. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Crystal structure of a human GABAA receptor". Nature. 512 (7514): 270–5. 2014. doi:10.1038/nature13293. PMC 4167603. PMID 24909990. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Perisynaptic localization of delta subunit-containing GABA(A) receptors and their activation by GABA spillover in the mouse dentate gyrus". J. Neurosci. 23 (33): 10650–61. November 2003. PMID 14627650. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Chapter 16: GABA and Glycine". Basic neurochemistry: molecular, cellular, and medical aspects (الطبعة Sixth). Philadelphia: Lippincott-Raven. 1999. ISBN 0-397-51820-X. مؤرشف من الأصل في 12 مايو 2009. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Role of GABAB receptors in GABA and baclofen-induced inhibition of adult rat cerebellar interpositus nucleus neurons in vitro". Brain Res Bull. 67 (4): 310–8. 2005. doi:10.1016/j.brainresbull.2005.07.004. PMID 16182939. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Yarom, M.; Tang, X. W.; Wu, E.; Carlson, R. G.; Vander Velde, D.; Lee, X.; Wu, J. (2016-08-01). "Identification of inosine as an endogenous modulator for the benzodiazepine binding site of the GABAA receptors". Journal of Biomedical Science. 5 (4): 274–280. ISSN 1021-7770. PMID 9691220. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Multiple facets of GABAergic neurons and synapses: multiple fates of GABA signalling in epilepsies". Trends Neurosci. 28 (2): 108–15. 2005. doi:10.1016/j.tins.2004.11.011. PMID 15667934. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Martin IL and Dunn SMJ. GABA receptors A review of GABA and the receptors to which it binds. Tocris Cookson LTD. نسخة محفوظة 22 يناير 2015 على موقع واي باك مشين.
- "Molecular composition of GABAC receptors". Vision Res. 38 (10): 1431–41. 1998. doi:10.1016/S0042-6989(97)00277-0. PMID 9667009. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Olsen, RW; Sieghart, W (January 2009). "GABA A receptors: subtypes provide diversity of function and pharmacology". Neuropharmacology. 56 (1): 141–8. doi:10.1016/j.neuropharm.2008.07.045. PMC 3525320. PMID 18760291. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Assembly and cell surface expression of heteromeric and homomeric g-aminobutyric acid type A receptors". J Biol Chem. 271 (1): 89–96. 1996. doi:10.1074/jbc.271.1.89. PMID 8550630. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Function and structure in glycine receptors and some of their relatives". Trends Neurosci. 27 (6): 337–44. 2004. doi:10.1016/j.tins.2004.04.010. PMID 15165738. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Haseneder, R; G, G; Zieglgänsberger, W; Kochs, E; Hapfelmeier, G (6 September 2002). "GABAA receptor activation and open-channel block by volatile anaesthetics: a new principle of receptor modulation?". European Journal of Pharmacology. 451 (1): 43–50. doi:10.1016/S0014-2999(02)02194-5. PMID 12223227. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Hunter, A (2006). "Kava (Piper methysticum) back in circulation". Australian Centre for Complementary Medicine. 25 (7): 529. الوسيط
|CitationClass=تم تجاهله (مساعدة) - (a) "Neurosteroid modulation of synaptic and extrasynaptic GABAA receptors". Pharmacology & Therapeutics. 116 (1): 20–34. 2007. doi:10.1016/j.pharmthera.2007.03.007. PMID 17531325. الوسيط
|CitationClass=تم تجاهله (مساعدة); (b) "Endogenous neurosteroids regulate GABAA receptors through two discrete transmembrane sites". Nature. 444 (7118): 486–9. 2006. doi:10.1038/nature05324. PMID 17108970. الوسيط|CitationClass=تم تجاهله (مساعدة); (c)"Characterization of brain neurons that express enzymes mediating neurosteroid biosynthesis". Proc. Natl. Acad. Sci. U.S.A. 103 (39): 14602–7. 2006. doi:10.1073/pnas.0606544103. PMC 1600006. PMID 16984997. الوسيط|CitationClass=تم تجاهله (مساعدة); (d) "Neurosteroid access to the GABAA receptor". J. Neurosci. 25 (50): 11605–13. 2005. doi:10.1523/JNEUROSCI.4173-05.2005. PMID 16354918. الوسيط|CitationClass=تم تجاهله (مساعدة); (e) "Neurosteroids: endogenous regulators of the GABAA receptor". Nat. Rev. Neurosci. 6 (7): 565–75. 2005. doi:10.1038/nrn1703. PMID 15959466. الوسيط|CitationClass=تم تجاهله (مساعدة); (f) "Fluoxetine and norfluoxetine stereospecifically and selectively increase brain neurosteroid content at doses that are inactive on 5-HT reuptake". Psychopharmacology (Berl.). 186 (3): 362–72. 2006. doi:10.1007/s00213-005-0213-2. PMID 16432684. الوسيط|CitationClass=تم تجاهله (مساعدة); (g) Dubrovsky BO (2005). "Steroids, neuroactive steroids and neurosteroids in psychopathology". Prog. Neuropsychopharmacol. Biol. Psychiatry. 29 (2): 169–92. doi:10.1016/j.pnpbp.2004.11.001. PMID 15694225. الوسيط|CitationClass=تم تجاهله (مساعدة); (h) "Neurosteroids: biochemistry and clinical significance". Trends Endocrinol. Metab. 13 (1): 35–43. 2002. doi:10.1016/S1043-2760(01)00503-3. PMID 11750861. الوسيط|CitationClass=تم تجاهله (مساعدة); (i) "Neurosteroids act on recombinant human GABAA receptors". Neuron. 4 (5): 759–65. 1990. doi:10.1016/0896-6273(90)90202-Q. PMID 2160838. الوسيط|CitationClass=تم تجاهله (مساعدة); (j) "Steroid hormone metabolites are barbiturate-like modulators of the GABA receptor". Science. 232 (4753): 1004–7. 1986. doi:10.1126/science.2422758. PMID 2422758. الوسيط|CitationClass=تم تجاهله (مساعدة); (k) "Neurosteroids — Endogenous Regulators of Seizure Susceptibility and Role in the Treatment of Epilepsy". Jasper's Basic Mechanisms of the Epilepsies [Internet]. 4th edition. Bethesda (MD): National Center for Biotechnology Information (US). 2012. مؤرشف من الأصل في 25 سبتمبر 2019. الوسيط|CitationClass=تم تجاهله (مساعدة) - Toraskar, Mrunmayee; Pratima R.P. Singh; Shashank Neve (2010). "STUDY OF GABAERGIC AGONISTS" (PDF). Deccan Journal of Pharmacology. 1 (2): 56–69. مؤرشف من الأصل (PDF) في 3 مارس 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Fisher JL (2009). "The anti-convulsant stiripentol acts directly on the GABA(A) receptor as a positive allosteric modulator". Neuropharmacology. 56 (1): 190–7. doi:10.1016/j.neuropharm.2008.06.004. PMC 2665930. PMID 18585399. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Boldyreva, A. A. (2005). "Lanthanum Potentiates GABA-Activated Currents in Rat Pyramidal Neurons of CA1 Hippocampal Field". Bulletin of Experimental Biology and Medicine. 140 (4): 403–5. doi:10.1007/s10517-005-0503-z. PMID 16671565. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Zinc-mediated inhibition of GABA(A) receptors: discrete binding sites underlie subtype specificity". Nat Neurosci. 6 (4): 362–9. 2003. doi:10.1038/nn1030. PMID 12640458. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "An inverse agonist selective for alpha5 subunit-containing GABAA receptors enhances cognition". J. Pharmacol. Exp. Ther. 316 (3): 1335–45. March 2006. doi:10.1124/jpet.105.092320. PMID 16326923. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "GABA A/Bz receptor subtypes as targets for selective drugs". Curr. Med. Chem. 14 (25): 2680–701. 2007. doi:10.2174/092986707782023190. PMID 17979718. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Affinity of 3-acyl substituted 4-quinolones at the benzodiazepine site of GABAA receptors". Bioorg. Med. Chem. 16 (14): 6936–48. July 2008. doi:10.1016/j.bmc.2008.05.049. PMID 18541432. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Differential organization of GABAA receptors and glycine receptors in the somatic and dendritic compartments of rat abducens motoneurons". Journal of Comparative Neurology. 504 (2): 112–126. 2007. doi:10.1002/cne.21442. PMID 17626281. مؤرشف من الأصل في 12 ديسمبر 2017. الوسيط
|CitationClass=تم تجاهله (مساعدة) - ten Hoeve AL (2012). GABA receptors and the immune system. Thesis, Utrecht University نسخة محفوظة 13 يونيو 2013 على موقع واي باك مشين.
مصادر خارجية
- Receptors, GABA-A في المَكتبة الوَطنية الأمريكية للطب نظام فهرسة المواضيع الطبية (MeSH).
- "Chapter 16: GABA and Glycine". Basic neurochemistry: molecular, cellular, and medical aspects (الطبعة Sixth). Philadelphia: Lippincott-Raven. 1999. ISBN 0-397-51820-X. مؤرشف من الأصل في 12 مايو 2009. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Chapter 16: GABA and Glycine". Basic Neurochemistry: Molecular, Cellular and Medical Aspects (الطبعة Seventh). Boston: Academic Press. 2005. صفحات 291–302. ISBN 0-12-088397-X. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Uusi-Oukari, M; Korpi, ER (2010). "Regulation of GABAA Receptor Subunit Expression by Pharmacological Agents". Pharmacological Reviews. 62 (1): 97–135. doi:10.1124/pr.109.002063. PMID 20123953. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Rudolph U (2015). Diversity and Functions of GABA Receptors: A Tribute to Hanns Möhler (الطبعة First). Academic Press, Elsevier. ISBN 978-0-12-802660-1. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علوم عصبية
بوابة علوم عصبية
