تنادد
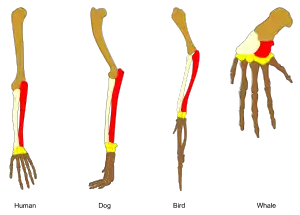
يشير مصطلح التنادد في علم الأحياء إلى وجود نسب مُشارَكِة بين زوج من البنى أو الجينات في أصناف مختلفة. أحد الأمثلة الشائعة على البنى المتناددة هي الأطراف الأماميّة في الفقاريات، حيث تكون أجنحة الخفَّاش و أذرع الرئيسيات و الزعانف الأماميّة للحيتان و الأرجل الأماميّة للكلاب و الأحصنة فجميعها مُشتَقَّة من بنية واحدة لدى رباعيات الأطراف التي تُمثِّلُ سلفاً مشتركاً لها جميعها. يشرح علم الأحياء التطوريّ فكرة التنادد بأن البنى المتناددة تكيَّفت (تأقلمت أو تلاءمت) لأغراض مختلفة كنتيجة للتوريث مع حدوث تعديل على المادة الوراثية للسلف المشترك. تم شرح التنادد بواسطة نظرية تشارلز داروين في التطوُّر عام 1859، ولكن تم ملاحظته قبل هذه النظريّة، من قبل أرسطو و من تبعه، وتم تحليلها بشكل واضح من قبل بيير بيلون في عام 1555. تم تطبيق هذا المصطلح homology في علم الأحياء من قبل عالم التشريح ريتشارد أوين عام 1843.
في علم الأحياء التطوُّريّ، تكون الأعضاء التي تطوَّرت في الجنين بنفس الطريقة ومن أصول مشابهة، كالمنشم المُطابق في القطاعات المتعاقبة لنفس الحيوان، تكون متناددة بشكل متسلسل. تتضمن أمثلة ذلك أرجل الحريشة، و اللامسة الفكيّة السفليّة واللامسة الشفويّة لدى الحشرات، و النواتئ الشوكيّة للفقرات المتعاقبة في العمود الفقريّ. الأعضاء التكاثريّة لدى الذكور والإناث متناددة إذ تطوَّروا من النسيج الجنينيّ نفسه، كما هو الحال بالنسبة للمبايض والخصى عند الثدييات بما فيها الإنسان.
يُعرف التنادد التسلسليّ بين البروتين و سلسلة دنا بشكل مماثل لقضية السلف المشترك. فشدفتان من دنا قد تتشاركان السلف لأنهما ناتجتين عن حادثة انتواع أو حادثة تكرار. يتم الاستدلال على التنادد بين البروتينات والدنا من خلال تشابه التسلسل. فالتشابه الواضح دليلٌ قويّ أن سلسلتين مرتبطتين بتطور متباعد من سلف مشترك. يُستخدم تراصف عدة تسلسلات لاكتشاف المناطق المتناددة.
يبقى التنادد مثيراً للجدل في سلوك الحيوان، ولكن هناك أدلة توحي بأن هرميّة الهيمنة على سبيل المثال متناددة بين الرئيسيات.
التاريخ
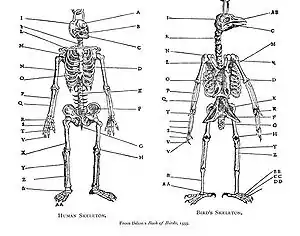
لاحظ أرسطو التنادد (350 قبل الميلاد) و حلَّله بيير بيلون في كتابه كتاب الطيور عام 1555، حيث قارن جهازياً بين هياكل الطيور والإنسان. و تم تفسير نموذج التشابه كجزء من سلسلة استقرار سلسلة الوجود العظمى خلال العصور الوسطى و أوائل العصر الحديث: و لم يُنظر إليه بعد ذلك كتغيُّرٍ تطوريّ. كما حاز التنادد أهميّة في التقاليد الطبيعيّة الفلسفيّة الألمانيّة، حيث نُظر إليه كتجلِّي لوحدة الطبيعة.[1][2] طرح جوته عام 17900 نظريته في مقال نشره بعنوان "ما وراء شكل النبات"، حيث أظهر خلال المقال أن أجزاء الزهرة مُشتَقَّةٌ من الأوراق.[3] وُصف التنادد المتسلسل للأطراف لاحقاً في القرن الثامن عشر. حيث أظهر عالم الحيوان الفرنسيّ إيتيان جوفري سانت هيلار عام 1818 في نظرية المتناددات أنه وُجد بُنىً متشارَكة بين الأسماك و الزواحف و الطيور و الثدييات. ذهب جوفري أبعد من ذلك، وسعى إلى إظهار التناددات بين تشعُّبات جورج كوفييه الواردة في كتاب مملكة الحيوان كالفقاريات و الرخويات، إلا أن ادعائه أثار نقاشات كوفييه-جوفري عام 1830. ذكر جوفري مبدأ الارتباطات، أي أن المهم هو الوضع النسبيّ للبنى المختلفة وارتباطاتها ببعضها البعض. ذكر عالم الجنين الإستونيّ كارل إرنست فون باير عام 1828 ما يُسمى حالياً بـقانون فون باير في 1828، حيث لاحظ أن الحيوانات المرتبطة تبدأ تطوُّرها كأجنة متماثلة ثم تفترق، لذا فإن حيوانات نفس الفصيلة أكثر قرباً وعندما تفترق تكون أكثر تأخُّراً في افترقها من افتراق الحيوانات الموجودة في الرتبة نفسها فقط (دون أن تكون في الفصيلة نفسها) و تمتلك متناددات أقل. تعترف نظرية فون باير أن كل أصنوفة (كالعائلة مثلاً) له خواص مُتشاركة مميَّزة، وأن مستويات التطوُّر الجنينيّ للهرميّة التصنيفيّة: ليست نفسها في نظريّة التجميع. استُخدم مصطلح "homology" "تنادد" لأول مرة في علم الأحياء من قبل عالم التشريح ريتشارد أوين عام 1843 عندما درس التشابهات بين زعانف وأطراف الفقاريات، مناقضاً لمصطلح "analogy" "التشابه" الذي كان يُستخدم لوصف البنى المختلفة ذات الوظيفة الواحدة. عام 1859 شرح تشارلز داروين البنى المتناددة على أنها تعني أن الكائنات المعنية تتشارك في خطة جسميّة بدءاً من سلف مشترك، وأن هذه الأصناف فروع لشجرة واحدة للحياة.[1][4][2]
التعريف
| ند [ مفرد ] : ج أنداد : مثل ونظير هو نده - لا ند له في السباق - تعامل مع فلان تعامل الند للند - { فلا تجعلوا لله أندادا } ° ما له ند : ما له نظير . |
| —معجم اللغة العربية المعاصرة |

صُكَّت كلمة homology حوالي عام 1656 مُشتَقَّةً من الكلمة الإغريقيّة (بالإغريقية: ὁμόλογος) التي تتألف من جذرين (بالإغريقية: ὁμός) و التي تعني "نفس" و (بالإغريقية: λόγος) و التي تعني "علاقة".[5][6][lower-alpha 1]
تتنادد البنى الأحيائيّة (البيولوجيّة) أو السلاسل في الأصنوفات المختلفة إذا كانت مُشتَقَّةً من سلف مشترك. فإن التنادد يعني تطور تباعدي. على سبيل المثال، العديد من الحشرات (كاليعسوب) تمتلك زوجين من أجنحة الطيران. في الخنافس، يتطور الزوج الأول من الأجنحة إلى أغطية جناحيّة قاسية (صناديق جناحيّة قاسية)،[9] بينما في ذوات الجناحين فإن الزوج الثاني من الأجنحة بتطور إلى أثقال صغيرة لتحقيق التوازن.[lower-alpha 2][10]
و بالمثل، فإن الأطراف الأمامية للسلف المشترك للفقاريات قد تطوَّرت إلى زعانف أمامية بالنسبة للحيتان و أجنحة بالنسبة للطيور و أرجل أماميّة للكلاب و الأيل و الأحصنة و إلى أرجل أماميّة قصيرة للضفادع و السحالي و إلى يدين بالنسبة للرئيسيات بما فيها الإنسان. جديرٌ بالذكر أن العظام الرئيسية للذراع (العضد و الكعبرة و الزند) موجودة في أحافير الأسماك لحمية الزعانف كأسماك Eusthenopteron.[11]
التنادد والتشابه
 مقالة مفصلة: تطور تقاربي
مقالة مفصلة: تطور تقاربي

يُعاكس مفهوم الأعضاء المتشابهة مفهوم الأعضاء المتناددة، حيث يكون للأعضاء المتشابهة وظائف متشابهة في أصنوفتين مختلفتين ولكن هذه الأعضاء لم تكن موجودةً في آخر سلف مشترك، و لكن بدلاً من ذلك تطوَّرا بشكل منفصل. على سبيل المثال، تطوَّرت أجنحة الطيور و الحشرات بشكل مستقل عن بعضهما البعض في مجموعات منفصلة على نطاق واسع، إلا أنها متقاربة وظيفياً لتُقدِّم وظيفة الطيران، لذا فإنها (أجنحة الطيور والحشرات) أعضاء متشابهة. و بالمثل، فإن أجنحة نبات القيقب الدلبي الكاذب و أجنحة الطيور أعضاءٌ متشابهة أيضاً ولكنها ليست متناددة، حيث أنهما (العضوان) يتطوران من بُنى مختلفة تماماً.[12][13] يمكن لأي بنية أن تنادد أخرى في مستوى واحد، وأن تشابه بنىً في مستوى آخر. فأجنحة التيروصور و الطيور و الخفافيش متشابهة كأجنحة وهي متناددة كأطراف أماميّة لأن العضو يخدم كذراع (و ليس كجناح) في آخر سلف مشترك لرباعيات الأطراف و تطوَّرت بطرق مختلفة في هذه المجموعات. لذا في التيروصور ينطوي الجناح على كونه من طرف أمامي وخلفيّ.[14] يُسمَّى التشابه homoplasy في الكلاديسيات أو التصنيفات التفرعيّة و (homoplasy) مصطلح يشير إلى تشارك الصفات من قِبل مجموعة من الأنواع الحيّة ولكنها (هذه الصفات غير موجودة في السلف المشترك. و من الأمثلة الجيّدة تطوُّر العين التي تنشأ بشكل مستقلّ في أنواع مختلفة، وعندما يحدث هذا يُدعى التقارب). في حين يُسمَّى في علم الأحياء التطوريّ بالتطوُّر التقاربيّ أو المتوازي.[15][16]
في التصنيفات التفرعية
- مقالة مفصلة: تصنيف تفرعي
تُستخدم المصطلحات التخصصيّة في بحوث التصنيف. فمصطلح التنادد الأوليّ (الأساسيّ) هو الفرضيَّةٌ الأوليّة التي يصوغها الباحث اعتماداً على بنية متشابهة أو اتصالات تشريحيّة مستنتجاً أن صفتين تتشاركان سلفاً. أما التنادد الثانويّ فيُضمَّن ضمن تحليل شحيح، حيث أن الصفة التي تحدث فقط مرَّةً على شجرة تؤخذ على أنها متناددة.[17] و كما هو وارد في التعريف، فإن العديد من التصانيف الفرعيّة (الكلاديستات) تعتبر التنادد ترادف مع الترادف الاشتقاقيّ الشكليّ، وهو صفة مُشتَقَّة مُتشاركة أو حالة خِلِّة (صفة) تُميِّز فرعاً حيوياً عن كائنات حية أخرى.[18]
في الأصنوفات المختلفة
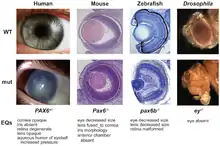
تقدِّمُ التناددات الأساس الأصوليّ للتصنيف الأحيائي (البيولوجيّ)، على الرغم من أن بعضها قد تكون غير بديهيّة (غير متوقَّعة) بشكل كبير. على سبيل المثال، التناددات العميقة كتناددات جينات PAX6 التي تتحكَّم بتطور العينين لدى الفقاريات و مفصليات الأرجل غير متوقَّعة، حيث تختلف الأعضاء تشريحياً وتبدو كما لو أنها قد تطوَّرت بشكل مستقلّ تماماً.[19][20]
في المفصليات
تباينت القطاعات الجسميّة الجنينيّة (الجسيدات) لمفصليات الأرجل المختلفة وتشعَّبت من مخطط جسميّ بسيط له عدة زوائد متماثلة، وهذه الزوائد متناددة بشكل تسلسليّ، وتطوَّر هذا المخطط البسيط إلى عدِّة مخططات جسميّة تمتلك شدفات ولها زوائد متخصِّصة.[21] و قد اِكتُشفت التناددات بين هذه البنى السابقة عبر مقارنة الجينات في ظل علم الأحياء التنمويّ التطوريّ.[19]

| جسيدة (قطعة الجسم) |
مفصليات ثلاثية الفصوص (الترايلوبيت) (مفصليات ثلاثية الفصوص) |
عنكبوت (كلابيات القرون)  |
حريشة (كثيرات الأرجل) |
حشرة (سداسيات الأرجل)  |
روبيان (قشريات) |
|---|---|---|---|---|---|
| 1 | قرون الاستشعار (هوائيات) | التآشير (الفك السفليّ والأنياب) | قرون الاستشعار (هوائيات) | قرون الاستشعار (هوائيات) | قرون الاستشعار (الهوائيات) الأولى |
| 2 | زوج الأرجل الأولى | لامسات | - | - | قرون الاستشعار (الهوائيات) الثانية |
| 3 | زوج الأرجل الثانية | زوج الأرجل الأولى | الأفكاك العلويّة | الأفكاك العلويّة | الأفكاك العلويّة |
| 4 | زوج الأرجل الثالثة | زوج الأرجل الثانية | الفك السفليّ الأول | الأفكاك السفليّة الأولى | الأفكاك السفليّة الأولى |
| 5 | زوج الأرجل الرابعة | الأرجل الثالثة | الأفكاك السفليّة الثانية | الأفكاك السفليّة الثانية | الأفكاك السفليّة الثانية |
| 6 | زوج الأرجل الخامسة | زوج الأرجل الرابعة | عنق (لا أرجل) | زوج الأرجل الأولى | زوج الأرجل الأولى |
| 7 | زوج الأرجل السادسة | - | زوج الأرجل الأولى | زوج الأرجل الثانية | زوج الأرجل الثانية |
| 8 | زوج الأرجل السابعة | - | زوج الأرجل الثانية | زوج الأرجل الثالثة | زوج الأرجل الثالثة |
| 9 | زوج الأرجل الثامنة | - | زوج الأرجل الثالثة | - | زوج الأرجل الرابعة |
| 10 | زوج الأرجل التاسعة | - | زوج الأرجل الرابعة | - | زوج الأرجل الخامسة |
بين الحشرات، يُمثِّلُ لاسع نحل العسل الإناث مسرأاً مُعدَّلاً، وهو منادد للمسرأ في الحشرات الأخرى كمستقيمات الأجنحة و نصفيات الأجنحة و غشائيات الأجنحة التي لا تحوي لاسعات.[22]
في الثدييات
- مقالة مفصلة: تشريح مقارن
تُستخدم عظيمات السمع الثلاث الموجودة في الأذن الوسطى لدى الثدييات بما فيها الإنسان، و هي المطرقة و السندان و الركاب، تُستخدم هذه العُظيمات الثلاث لنقل الصوت من غشاء الطبل إلى الأذن الداخليّة. تطوَّرت عظام المطرقة والسندان في الجنين من بُنى تُشكِّلُ عظام الفك (العظام المُربَّع والمِفصليّ) في السحالي وفي الأحافير الشبيهة بالسحالي من أسلاف الثدييات. كلا خطي الأدلة يُظهران أن هذه العظيمات الثلاث مُناددة وتتشارك سلفاً مشتركاً.[23]
و من بين العديد من المتناددات في الأجهزة التكاثريّة للثدييات، تبرز المبايض و الخصى كأبرز المتناددات.[24]
يمكن فهم الأعضاء البدائيّة كعظم العصعص (الذيل البشريّ)، و التي تقلّ الآن كثيراً عن حالتها الوظيفيّة، يمكن فهمها كإشارات للتطوُّر، حيث تُفسَّر هذه الإشارات على أن الانتخاب الطبيعيّ تخفيض هذه الأعضاء من أجهزتها الوظيفيّة عندما لم تعد هناك حاجة لوظائفها، ولا يوجد أي معنى على الإطلاق لوجودها إذا ما كانت الأنواع الحيّة ثابتة. فعظم العصعص لدى الإنسان منادد للذيل لدى الرئيسيات الأخرى.[25]
في النباتات
الأوراق والجذوع و الجذور
في العديد من النباتات، تُصنَع بُنى دفاعيّة أو تخزينيّة عبر تعديل تطوُّر الأوراق أو الساق أو الجذور الأوليّة. تُعدَّل الأوراق بشكل مختلف من بُنى التركيب الضوئي لتُشكِّل مادة مُلصِقة على سطوحها للحشرات بالنسبة لنبات الإبريق، و فكين مُحاصرين لخناق الذباب، وأشواكاً بالنسبة للصبار، و جميع هذه البنى (الأباريق الهاضمة للحشرات والفكين المحاصرين وأشواك الصبَّار) متناددة.[26]
| الأعضاء الأوليّة | البنى الدفاعيّة | البنى التخزينيّة |
|---|---|---|
| الأوراق | الأشواك | الأوراق المتورِّمة (مثلاً النباتات العصارية) |
| الجذوع | الإبر | الدرنات (مثلاً البطاطا) و الجذمور (مثلاً الزنجبيل) و الجذوع اللحميّة (الصبَّار) |
| الجذور | - | الدرنات الجذريّة (مثلاً البطاطا الحلوة) و الجذور الوتديّة (مثلاً الجزر) |
تتنادد أوراق مُركَّبة محددة لنباتات مُزهرة مع الأوراق والبراعم على حدٍّ سواء، لأن تناميها قد تطوَّر من فسيفساء جينيّة لتطوُّر أوراق وبراعم.[27][28]
 ورقة ريشيّة واحدة لنبات المران العالي
ورقة ريشيّة واحدة لنبات المران العالي.jpg.webp) تفصيل لورقَة راحيّة
تفصيل لورقَة راحيّة أوراق سويقات تأقلمت (تكيَّفت لتكون) كأشواك في نبات Fouquieria splendens
أوراق سويقات تأقلمت (تكيَّفت لتكون) كأشواك في نبات Fouquieria splendens الأوراق الكبيرة جداً للموز، موز مستدق
الأوراق الكبيرة جداً للموز، موز مستدق ورقة واذمة مائيّة تخزينيّة لنبات الصبر
ورقة واذمة مائيّة تخزينيّة لنبات الصبر ورقة مُحاصرة للحشرات من نبات خناق الذباب
ورقة مُحاصرة للحشرات من نبات خناق الذباب ورقة مُحاصرة للحشرات تعود لنبات الإبريق
ورقة مُحاصرة للحشرات تعود لنبات الإبريق
أجزاء الزهرة
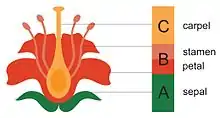
تتنادد الأنماط الأربعة لأجزاء الزهرة وهي المتاع (المدقة) و الأسدية و البتلات و كأس الزهرة (السبلات)، و تُشتَقُّ من الأوراق كما لاحظ جوته في عام 1790. وصف جوته كذلك تنامي هذه الأجزاء خلال نموذج من نماذج التعبير الجينيّ في المناطق الناميّة (البارضات) بنموذج أ ب ج لتنامي الزهرة. يتنامى كل نمط من الأنماط الأربعة لأجزاء الزهرة بشكل متسلسل في إطار حلقات عمل مُعيَّنة، تتحكَّم بها عدد قليل من الجينات التي تمارس تأثيراً بأجزاء زهريّة مختلفة غالباً. لذا فإن الجينات أ تعمل وحيدةً مما يؤدي لتشكيل كأس الزهرة (السبلات)، بينما تعمل الجينات أ وب معاً لتشكيل البتلات، وتعمل الجينات ب وج نعاً لخلق الأسدية، فيما تعمل الجينات ج وحيدةً لتشكيل المتاع (المدقة). و عندما لا يكون أي مجموعة من مجموعات الجينات السابقة مُفعَّلةً، تتشكَّل الأوراق. هناك أيضاً مجموعتان جينيَّتان أخريان هما المجموعة د لتشكيل البويضات والمجموعة هـ (E) للتجعُّدات الزهريّة. هذه الجينات قديمة بشكل واضح، كقدم النباتات المزهرة بحد ذاتها.[3]
علم الأحياء التطوري

يمكن لعلم الأحياء التطوري أن يحدد البنى المتناددة التي تنشأ من النسيج نفسه في التخلُّق الجنينيّ. على سبيل المثال، لا تمتلك الثعابين البالغة أرجلاً، ولكن لأجنتها في مراحل باكرة براعم طرفيّة لأرجل خلفيّة، ولكنها تتراجع بتنامي الجنين. يؤكَّد امتلاك أسلاف الثعابين لأرجلٍ خلفيّة من خلال الأدلة الأحفوريّة: يمتلك الثعبان الطباشيريّ Pachyrhachis problematicus أرجلاً خلفيّة كاملة تشتمل على ورك (عظام الحرقفة و العانة و الإسك) و عظم فخذيّ (فخذ) و عظمي ساق (ظنبوب و شظية) و عظام أقدام (عظم العَقِب و عظم الكاحل) كما هو الحال الآن في رباعيات الأطراف.[29]
التنادد التسلسلي
- مقالات مفصلة: تنادد عميق
- علم الأحياء النمائي التطوري

كما هو الحال في البُنى التشريحيّة، يُعرَّف التنادد التسلسليّ بين البروتين أو سلاسل الدنا بأنه الاشتراك بسلف. يمكن أن يتشارك تسلسلان لدنا بسلف لأنهما ناجمان عن حادثة انتواع أو تضاعف. عادةً ما يُستدَلُّ على التنادد بين البروتينات والدنا من تشابه التسلسلات. و التماثل الواضح دليلٌ قويّ أن تسلسلان مرتبطان بتطوُّر متباعد لسلف مشترك. يُستخدم ترصيف عدة سلاسل للإشارة إلى أيٍّ من المناطق من كل سلسلة متناددة.[31]
تكون التسلسلات المتناددة ناجمة عن حادثة انتواع إذا انحدرت جميعها من تسلسل سلف ذاته ويحدث الانفصال بحادثة انتواع: عندما ينفصل نوعٌ ما إلى نوعين منفصلين، يُقال حينها عن نسخ الجين الموجودة في النوعين الناتجين، يُقال عنها أنها متناددة ناجمة عن حادثة انتواع. تمت صياغة مُصطلح ortholog و orthologous فيما بعد للإشارة إلى التنادد الناجم عن انتواع، وصِيغ هذا المُصطلح عام 1970 من قِبل عالم التطوُّر الجزيئيّ والتر فيتش.[32]
بينما يُشار إلى التسلسلات المتناددة بِكلمة paralogous عند الحديث عن تسلسلان متناددان ناجمان عن حادثة تضاعف داخل الجينوم. و بالنسبة لأحداث التضاعف الجينيّة، إذا تضاعف جين داخل كائن حيّ ليحتلَّ موقعين في الجينوم ذاته، يُطلق على الجينين النسختين صفة paralogous. تنتمي الجينات الناجمة عن حادثة تضاعف غالباً إلى الأنواع ذاتها. و من الممكن أن تشكل شكل بنية الجينوم بأكمله وبالتالي تشرح تطوُّر الجينوم إلى محتوى كبير. تتضمن الأمثلة، العلبة المثلية أو ما يُدعى بصندوق النحَّت (جينات النحَّت) في الحيوانات. لاتخضع هذه الجينات إلى التضاعف الجينيّ داخل الصبغيات، بل تضاعفات الجينوم بأكلمه. و كنتيجة، تنتشر جينات النحَّت في معظم الفقاريات بين صبغيات متعددة: و قد دُرست تجمُّعات النحَّت A-D بشكل جيِّد في هذا المجال.[33]
.JPG.webp)
في السلوك
اقترح أن بعض السلوكيَّات قد تتنادد، اعتماداً على التشارك بين الأصنوفات المرتبطة أو على الأصول الشائعة لهذه السلوكيَّات في تطوُّر الأفراد، على الرغم من أن هذا الطرح ما يزال مُثيراً للجدل.[34] على سبيل المثال، يجادل دي. دبليو. راجيكي وراندال سي. فلانيري، مستخدمين بيانات متعلِّقة بالبشر والرئيسيات مما سوى البشر، يجادلان بأن نماذج السوك في هرميَّات الهيمنة متناددة بين الرئيسيّات.[35]
التنادد وعلم الوراثة
هناك فكرةٌ تنص على احتمال وجود المعلومات بشكل مشفرٍ في جينات الكائن الحي. في ثلاثينيات القرن العشرين، فسّر تخليق نظرية داروين وعلم الوراثة السكانية التطورَ باعتباره تغييراً في ترددات الجينات، وبعد عدة عقود أوصل اكتشاف بنية ووظيفة الـ DNA هذا التفسير إلى المستوى الجزيئي.
الاستنتاج
في عام 1802، كتب ويليام بيلي مثالاً عن شخص ما يعبر أرضاً خضراء ويجد حجراً، يمكن لهذا الشخص أن يعزو وجود الحجر لأسباب طبيعية غير هادفة. أما عند العثور على ساعة مثلاً، سيلاحظ الشخص وجود “عدة أجزاء مؤطرة ومجمعة لغرض معين”. يستنتج المرء في هذه الحالة أن الساعة قد تم تصميمها. من خلال التجانس، يستنتج المرء أيضاً أن الكائنات الحية هي الأخرى مصممة. في عام 1859، ادعى تشارلز داروين أن الكائنات الحية تشبه حجر بيلي أكثر من الساعة، وادعى أن كل شيء نسبه بيلي إلى التصميم يمكن أن يُعزى بشكل طبيعي من خلال الأصل والتعديل.[36]
تمتلك الكائنات المتنوعة سمات متجانسة. وقد يكون (أو لا يكون) التنادد في بعض الحالات بسبب التوارث من سلف مشترك، لكنه بالتأكيد لا يرجع إلى تشابه الجينات أو تشابه مسارات النمو. في عام 1971، كتب غافن دي بير التالي: “ما الآلية التي يمكن أن تؤدي إلى إنتاج أعضاء متجانسة، نفس ”الأنماط “، حتى إن لم تسيطر الجينات عليها؟ لقد طرحت هذا السؤال في عام 1938، ولم يرد أحد عليه”. بعد مرور ستة وعشرين عاماً، لمّا يجب أحدٌ على هذا السؤال.
ملاحظات
- استخدمت أيضاً المصطلحات البديلة"homogeny" "التجانس" و "homogenous" و "المتجانس" في آواخر القرن التاسع عشر و أوائل القرن العشرين. على أي حال، فإن هذه المصطلحات قديمة في علم الأحياء و مصطلح "homogenous" يترافق حالياً كخطأ إملائي مع المصطلح "homogeneous" الذي يشير إلى تماثل من الخليط.[7][8]
- إذا تم اعتبار زوجي الأجنحة بنى متناددة و قابلة للخضوع للتغيرات، فإن هذا قد يُوصف كتخفيض متوازٍ في عدد الأجنحة، و لكن على خلاف ذلك يكون التغير الطارئ على الزوجين تغيراً متباعداً (لا علاقة للتغييرات الحاصلة على الزوج الأول بالتغيرات الحاصلة على الزوج الثاني).
مراجع
- Panchen, A. L. (1999). "Homology—history of a concept". Novartis Found Symp (222): 5–18. PMID 10332750. مؤرشف من الأصل في 29 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Brigandt, Ingo (23 November 2011). "Essay: Homology". The Embryo Project Encyclopedia. مؤرشف من الأصل في 11 مارس 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Dornelas, Marcelo Carnier; Dornelas, Odair (2005). "From leaf to flower: Revisiting Goethe's concepts on the ¨metamorphosis¨ of plants". Brazilian Journal of Plant Physiology. 17 (4). doi:10.1590/S1677-04202005000400001. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Sommer, R. J. (July 2008). "Homology and the hierarchy of biological systems". BioEssays. 30 (7): 653–658. doi:10.1002/bies.20776. PMID 18536034. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bower, Frederick Orpen (1906). "Plant Morphology". Congress of Arts and Science: Universal Exposition, St. Louis, 1904. Houghton, Mifflin. صفحة 64. مؤرشف من الأصل في 14 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Williams, David Malcolm; Forey, Peter L. (2004). Milestones in Systematics. CRC Press. صفحة 198. ISBN 0-415-28032-X. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "homogeneous, adj.". OED Online. March 2016. Oxford University Press. http://www.oed.com/view/Entry/88045? (accessed April 09, 2016). نسخة محفوظة 23 نوفمبر 2020 على موقع واي باك مشين.
- "homogenous, adj.". OED Online. March 2016. Oxford University Press. http://www.oed.com/view/Entry/88055? (accessed April 09, 2016). نسخة محفوظة 3 ديسمبر 2014 على موقع واي باك مشين.
- Wagner, Günter P. (2014). Homology, Genes, and Evolutionary Innovation. Princeton University Press. صفحات 53–54. ISBN 978-1-4008-5146-1. مؤرشف من الأصل في 13 ديسمبر 2019.
elytra have very little similarity with typical wings, but are clearly homologous to forewings. Hence butterflies, flies, and beetles all have two pairs of dorsal appendages that are homologous among species.
الوسيط|CitationClass=تم تجاهله (مساعدة) - Lipshitz, Howard D. (2012). Genes, Development and Cancer: The Life and Work of Edward B. Lewis. Springer. صفحة 240. ISBN 978-1-4419-8981-9. مؤرشف من الأصل في 23 يناير 2020.
For example, wing and haltere are homologous, yet widely divergent, organs that normally arise as dorsal appendages of the second thoracic (T2) and third thoracic (T3) segments, respectively.
الوسيط|CitationClass=تم تجاهله (مساعدة) - "Homology: Legs and Limbs". UC Berkeley. مؤرشف من الأصل في 30 يناير 2018. اطلع عليه بتاريخ 15 ديسمبر 2016. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Secret Found to Flight of 'Helicopter Seeds'". LiveScience. 11 June 2009. مؤرشف من الأصل في 28 يونيو 2018. اطلع عليه بتاريخ 02 مارس 2017. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lentink, D.; Dickson, W. B.; van Leeuwen, J. L.; Dickinson, M. H. (12 Jun 2009). "Leading-Edge Vortices Elevate Lift of Autorotating Plant Seeds". Science. 324 (5933): 1438–1440. doi:10.1126/science.1174196. الوسيط
|CitationClass=تم تجاهله (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Scotland, R. W. (2010). "Deep homology: A view from systematics". BioEssays. 32 (5): 438–449. doi:10.1002/bies.200900175. PMID 20394064. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cf. Butler, A. B.: Homology and Homoplasty. In: Squire, Larry R. (Ed.): Encyclopedia of Neuroscience, Academic Press, 2009, pp. 1195–1199.
- "Homologous structure vs. analogous structure: What is the difference?". مؤرشف من الأصل في 17 أغسطس 2017. اطلع عليه بتاريخ 27 سبتمبر 2016. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Pinna, M. C. C. (1991). "Concepts and Tests of Homology in the Cladistic Paradigm". Cladistics. 7 (4): 367–394. doi:10.1111/j.1096-0031.1991.tb00045.x. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Page, Roderick D.M.; Holmes, Edward C. (2009). Molecular Evolution: A Phylogenetic Approach. John Wiley & Sons. ISBN 978-1-4443-1336-9. مؤرشف من الأصل في 14 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Brusca, R.C.; Brusca, G.J. (1990). Invertebrates. Sinauer Associates. صفحة 669. الوسيط
|CitationClass=تم تجاهله (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Carroll, Sean B. (2006). Endless Forms Most Beautiful. Weidenfeld & Nicolson. صفحات 28, 66–69. ISBN 0-297-85094-6. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Novartis Foundation; Hall, Brian (2008). Homology. John Wiley. صفحة 29. ISBN 978-0-470-51566-2. مؤرشف من الأصل في 24 يناير 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Shing, H.; Erickson, E. H. (1982). "Some ultrastructure of the honeybee (Apis mellifera L.) sting". Apidologie. 13 (3): 203–213. مؤرشف من الأصل في 15 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Homology: From jaws to ears — an unusual example of a homology". UC Berkeley. مؤرشف من الأصل في 30 يناير 2018. اطلع عليه بتاريخ 15 ديسمبر 2016. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Hyde, Janet Shibley; DeLamater, John D. (June 2010), "Chapter 5" (PDF), Understanding Human Sexuality (الطبعة 11th), New York: ماكجرو هيل التعليم, صفحة 103, ISBN 978-0-07-338282-1 الوسيط
|CitationClass=تم تجاهله (مساعدة); الوسيط|separator=تم تجاهله (مساعدة)CS1 maint: ref=harv (link) - Larson 2004، صفحة 112.
- "Homology: Leave it to the plants". University of California at Berkeley. مؤرشف من الأصل في 30 يناير 2018. اطلع عليه بتاريخ 07 مايو 2017. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Sattler R (1984). "Homology — a continuing challenge". Systematic Botany. 9 (4): 382–94. doi:10.2307/2418787. JSTOR 2418787. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Sattler, R. (1994). "Homology, homeosis, and process morphology in plants". In Hall, Brian Keith (المحرر). Homology: the hierarchical basis of comparative biology. Academic Press. صفحات 423–75. ISBN 0-12-319583-7. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Homologies: developmental biology". UC Berkeley. مؤرشف من الأصل في 02 أكتوبر 2017. اطلع عليه بتاريخ 15 ديسمبر 2016. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ أرشيف=(مساعدة) - "Clustal FAQ #Symbols". Clustal. مؤرشف من الأصل في 06 يوليو 2018. اطلع عليه بتاريخ 08 ديسمبر 2014. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ أرشيف=(مساعدة) - Koonin EV (2005). "Orthologs, paralogs, and evolutionary genomics". Annual Review of Genetics. 39: 309–38. doi:10.1146/annurev.genet.39.073003.114725. PMID 16285863. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Fitch, W.M. (June 1970). "Distinguishing homologous from analogous proteins". Systematic Zoology. 19 (2): 99–113. doi:10.2307/2412448. JSTOR 2412448. PMID 5449325. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Zakany, Jozsef; Duboule, Denis (2007). "The role of Hox genes during vertebrate limb development". Current Opinion in Genetics & Development. 17 (4): 359–366. doi:10.1016/j.gde.2007.05.011. ISSN 0959-437X. PMID 17644373. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Moore, David S (2013). "Importing the homology concept from biology into developmental psychology". Developmental Psychobiology. 55 (1): 13–21. doi:10.1002/dev.21015. PMID 22711075. مؤرشف من الأصل في 17 أغسطس 2017. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Rajecki, D. W.; Flanery, Randall C. (2013). Lamb, M. E.; Brown, A. L. (المحرر). Social Conflict and Dominance in Children: a Case for a Primate Homology. Advances in Developmental Psychology. Taylor and Francis. صفحة 125. ISBN 978-1-135-83123-3. مؤرشف من الأصل في 12 ديسمبر 2019.
Finally, much recent information on children's and nonhuman primates' behavior in groups, a conjunction of hard human data and hard nonhuman primate data, lends credence to our comparison. Our conclusion is that, based on their agreement in several unusual characteristics, dominance patterns are homologous in primates. This agreement of unusual characteristics is found at several levels, including fine motor movement, gross motor movement, and behavior at the group level.
الوسيط|CitationClass=تم تجاهله (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Homology-history of a concept. - PubMed - NCBI نسخة محفوظة 01 أبريل 2019 على موقع واي باك مشين.
 بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء التطوري
بوابة علم الأحياء التطوري
 صور وملفات صوتية من كومنز
صور وملفات صوتية من كومنز
