أكوابورين
أكوابورين الأكوابورينات (بالإنجليزية: Aquaporin)، وتسمى أيضًا قنوات الماء، هي بروتينات من عائلة كبيرة من البروتينات الداخلية الرئيسية التي تشكل مسامًا في غشاء الخلايا الحية، مما يسهل نقل الماء بين الخلايا بشكل أساسي.[1] تحتوي أغشية الخلايا لمجموعة متنوعة من البكتيريا والفطريات والخلايا الحيوانية والنباتية على أكوابورينات يمكن للمياه أن تتدفق من خلالها بسرعة أكبر داخل وخارج الخلية أكثر من الانتشار عبر طبقة ثنائية الفوسفوليبيد (غشاء الخلية).[2] تحتوي الأكوابورينات على ستة مجالات حلزونية ألفا تغطي الأغشية مع كل من النهايات الكربوكسيلية والأمينية على الجانب السيتوبلازمي. تحتوي حلقتان كارهتان للماء على شكل NPA محفوظ من الأسباراجين والبرولين والألانين والذي يشكل برميلًا يحيط بمنطقة تشبه المسام المركزية التي تحتوي على كثافة بروتينية إضافية. نظرًا لأن الأكوابورينات عادةً ما تكون مفتوحة دائمًا ومنتشرة في كل نوع من الخلايا تقريبًا، فإن هذا يؤدي إلى سوء فهم مفاده أن الماء يمر بسهولة عبر غشاء الخلية تحت تدرج تركيزه. يمكن أن يمر الماء عبر غشاء الخلية من خلال الانتشار البسيط لأنه جزيء صغير، ومن خلال التناضح، في الحالات التي يكون فيها تركيز الماء خارج الخلية أكبر من تركيزه داخلها. ومع ذلك، نظرًا لأن الماء هو جزيء قطبي، فإن عملية الانتشار البسيطة هذه بطيئة نسبيًا، وتمر غالبية الماء عبر الأكوابورين.[3][4][5]
| Aquaporin | |
|---|---|
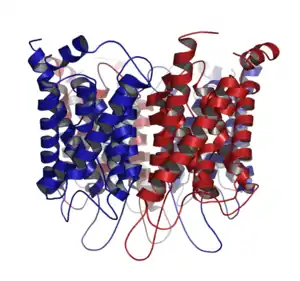 | |
| التركيب البلوري للأكوابورين 1 | |
| معرف | |
| رمز | Aquaporin |
| قاعدة بيانات عوائل البروتينات | PF00230 |
| إنتربرو | IPR000425 |
| بروسايت | PDOC00193 |
| قاعدة بيانات التصنيف الهيكلي للبروتينات | 1fx8 |
| قاعدة بيانات تصنيف الناقل [الفرنسية] | 1.A.8 |
| قاعدة بيانات توجهات البروتينات في الأغشية | 7 |
| قاعدة بيانات توجهات البروتينات في الأغشية | 2zz9 |
مُنحت جائزة نوبل في الكيمياء لعام 2003 مناصفةً لكل من "بيتر أجري" لاكتشاف الأكوابورينات[6] و"رودريك ماكينون" عن عمله على تركيب وآلية عمل قنوات البوتاسيوم.[7]
ارتبطت العيوب الجينية التي تنطوي على جينات الأكوابورين بالعديد من الأمراض البشرية بما في ذلك مرض السكري الكاذب الكلوي والتهاب النخاع والعصب البصري.[8][9][10][11]
مقدمة
يمكن أن ينتشر الماء بحرية من خلال طبقة ثنائية الدهون بمعدل محدود، ولكن لوحظ أن نفاذية الماء في خلايا الدم الحمراء كانت أعلى بكثير مما كان متوقعًا للانتشار التلقائي. دفعت هذه الملاحظة إلى البحث عن بروتينات متخصصة قد تكون موجودة لتسهيل تدفق المياه عبر الأغشية. تم تنقية قناة الماء المغلفة من غشاء خلايا الدم الحمراء في البداية باعتبارها نوعًا مرتبطًا بغشاء 28 كيلو دالتون، يُطلق عليه اسم بروتين متكامل مكون من 28 كيلو دالتون (CHIP28) (Denker et al. 1988). تم عرض التعبير عن CHIP28 inXenopusoocytes لاحقًا لإحداث انتشار سريع للمياه أسفل التدرجات التناضحية عبر غشاء البلازما، مما يؤدي إلى تحديد أول قناة موصلة للماء (Prestonet al. 1992). اقترح تحليل تسلسل البروتين أن CHIP28 متماثل مع البروتين الداخلي الرئيسي (MIP) من خلايا ألياف العدسة (جورين وآخرون 1984)، وبعد وقت قصير من إنشاء وظيفة مسام الماء في CHIP28، تم العثور على البروتينات المتماثلة في الثدييات والبكتيريا والنباتات تعمل كقنوات مائية، ومصطلح "أكوابورين" تم صياغته لأعضاء عائلة البروتين هذه (أجري، ساساكي وكريسبيلز 1993)، وبناءً عليه، تمت إعادة تسمية CHIP28 إلى AQP1 وأصبح MIP يُعرف الآن باسم AQP0.
التاريخ
جذبت آلية النقل المائي الميسر والوجود المحتمل لمسام المياه الباحثين منذ عام 1957.[12] في معظم الخلايا، ينتقل الماء للداخل والخارج عن طريق التناضح من خلال المكون الدهني لأغشية الخلايا. نظرًا لارتفاع نفاذية الماء نسبيًا لبعض الخلايا الظهارية، كان يشتبه منذ فترة طويلة في وجود آلية إضافية لنقل المياه عبر الأغشية. قام سولومون وزملاؤه بعمل رائد في مجال نفاذية الماء عبر غشاء الخلية في أواخر الخمسينيات من القرن الماضي. في منتصف الستينيات، سعت فرضية بديلة ("نموذج التقسيم-الانتشار") إلى إثبات أن جزيئات الماء تنقسم بين طور الماء ومرحلة الدهون ثم تنتشر عبر الغشاء، وتعبرها حتى الطور البيني التالي حيث تترك[13][14] وعاد الدهن إلى الطور المائي. دراسات باريسي، إيدلمان، كارفونيس وآخرون. أكد ليس فقط على أهمية وجود القنوات المائية ولكن أيضًا على إمكانية تنظيم خصائص نفاذيتها.[15][16] في عام 1990، أظهرت تجارب فيركمان التعبير الوظيفي للقنوات المائية، مما يشير إلى أن قنوات المياه عبارة عن بروتينات فعالة.[17][18][19][20][21]
الاكتشاف
أُبلغ عن أول أكوابورين، "أكوابورين -1" (المعروف أصلاً باسم CHIP 28)، بواسطة بيتر أجري، من جامعة جونز هوبكنز عام 1992.[22] في عام 1999، مع فرق بحثية أخرى، قدم "أجري" أول صور عالية الدقة للبنية ثلاثية الأبعاد للأكوابورين، وهي أكوابورين -1. حددت دراسات أخرى باستخدام محاكاة الكمبيوتر العملاق مسار الماء أثناء تحركه عبر القناة وأظهرت كيف يمكن للمسام أن تسمح بمرور الماء دون مرور المواد المذابة الصغيرة. حصل البحث الرائد والاكتشاف اللاحق لقنوات المياه من قبل أجري وزملائه على جائزة نوبل في الكيمياء عام 2003.[23] قال أجري إنه اكتشف الأكوابورينات "بالصدفة". كان يدرس مستضدات فصيلة الدم Rh وعزل جزيء Rh، لكن الجزيء الثاني، بحجم 28 كيلو دالتون (وبالتالي يُسمى 28K) استمر في الظهور. في البداية ظنوا أنه جزء من جزيء Rh، أو ملوث، لكن اتضح أنه نوع جديد من الجزيئات ذات وظيفة غير معروفة. كانت موجودة في هياكل مثل الأنابيب الكلوية وخلايا الدم الحمراء، وترتبط ببروتينات من أصول متنوعة، مثل دماغ ذبابة الفاكهة والبكتيريا وعدسة العين وأنسجة النبات.
ومع ذلك، فإن التقرير الأول عن نقل المياه عبر الأغشية بواسطة البروتين كان بقلم جورج بنجا وآخرون في عام 1986، قبل نشر أجري الأول حول هذا الموضوع.[24] أدى ذلك إلى جدل مفاده أن عمل بنجا لم يتم الاعتراف به بشكل كاف سواء من قبل أجري أو من قبل لجنة جائزة نوبل.[25][26][27]
الوظيفة
الأكوابورين هو "نظام تصريف المياه للخلايا". ينتقل الماء عبر الخلايا بطريقة منظمة، وبسرعة أكبر في الأنسجة التي تحتوي على قنوات مائية للأكوابورين. لسنوات عديدة، افترض العلماء أن الماء يتسرب عبر غشاء الخلية، وبعض الماء يفعل. ومع ذلك، فإن هذا لم يفسر كيف يمكن للماء أن يتحرك بهذه السرعة عبر بعض الخلايا.[28]
تقوم الأكوابورينات بشكل انتقائي بتوصيل جزيئات الماء داخل وخارج الخلية، مع منع مرور الأيونات والمواد المذابة الأخرى. تُعرف الأكوابورينات أيضًا باسم قنوات المياه، وهي بروتينات غشائية متكاملة. بعضها، المعروف باسم aquaglyceroporins، ينقل أيضًا جزيئات أخرى صغيرة مذابة غير مشحونة بما في ذلك الأمونيا وثاني أكسيد الكربون والجلسرين واليوريا. على سبيل المثال، قناة aquaporin 3 لها عرض مسامي 8-10 أنغستروم وتسمح بمرور الجزيئات المحبة للماء تتراوح بين 150 و200 دا. ومع ذلك، فإن مسام الماء تسد الأيونات تمامًا بما في ذلك البروتونات، وهي ضرورية للحفاظ على فرق الجهد الكهروكيميائي للغشاء.[29] تعبر جزيئات الماء عبر مسام القناة في ملف واحد. يزيد وجود قنوات مائية من نفاذية الغشاء للماء. وهي أيضًا ضرورية لنظام نقل المياه في النباتات وتحمل الجفاف وضغوط الملح.
التركيب
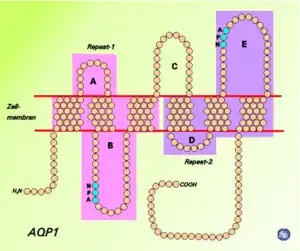
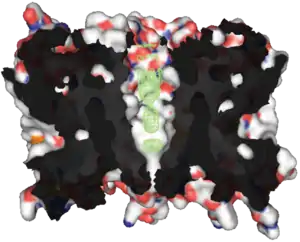
تتكون بروتينات Aquaporin من حزمة من ستة α-helices عبر الغشاء. يتم تضمينها في غشاء الخلية. تواجه النهايات الأمينية والكربوكسيلية داخل الخلية.[30] يتشابه نصفا الأمينو والكربوكسيل مع بعضهما البعض، ويبدو أنهما يكرران نمطًا من النيوكليوتيدات. يعتقد بعض الباحثين [من؟] أن هذا تم إنشاؤه من خلال مضاعفة جين نصف الحجم سابقًا. يوجد بين الحلزونات خمس مناطق (A - E) تدور داخل أو خارج غشاء الخلية، اثنتان منها كارهة للماء (B، E)، مع نمط أسباراجين - برولين - ألانين ("عزر NPA"). إنها تخلق شكل الساعة الرملية المميز، مما يجعل قناة المياه ضيقة في المنتصف وأوسع عند كل طرف.
مكان آخر أضيق في القناة هو "مرشح انتقائية ar / R"، وهو مجموعة من الأحماض الأمينية تمكن الأكوابورين من السماح بمرور جزيئات مختلفة أو منع مرورها بشكل انتقائي.
تشكل الأكوابورينات عناقيد من أربعة أجزاء (رباعيات) في غشاء الخلية، حيث يعمل كل واحد من المونومرات الأربعة كقناة مائية. تحتوي الأكوابورينات المختلفة على قنوات مائية مختلفة الأحجام، وهي أصغر الأنواع لا تسمح بمرور أي شيء سوى الماء.
تظهر ملامح الأشعة السينية أن الأكوابورينات لها مدخلين مخروطي الشكل. يمكن أن يكون شكل الساعة الرملية هذا نتيجة لعملية اختيار طبيعية نحو النفاذية المثلى. [بحث أصلي؟] وقد ثبت أن المداخل المخروطية بزاوية فتح مناسبة يمكن أن توفر بالفعل زيادة كبيرة في نفاذية القناة الهيدروديناميكية.
NPA motif
تظهر قنوات Aquaporin في عمليات المحاكاة للسماح بمرور الماء فقط، حيث تصطف الجزيئات بشكل فعال في ملف واحد. وبتوجيه من المجال الكهربائي المحلي للأكوابورين، يتجه الأكسجين الموجود في كل جزيء ماء للأمام أثناء دخوله، ويستدير في منتصف الطريق تقريبًا ويترك الأكسجين متجهًا للخلف. لماذا يحدث هذا الدوران ليس واضحًا تمامًا بعد. حدد بعض الباحثين مجالًا إلكتروستاتيكيًا ناتجًا عن نصف حلزونات أكوابورين HB وHE كسبب. اقترح آخرون أن الروابط الهيدروجينية بين الأحماض الأمينية الأسباراجين في منطقتي NPA والأكسجين في الماء يسبب الدوران. لا يزال من غير الواضح ما إذا كان دوران جزيئات الماء له أي أهمية بيولوجية. تكهنت الدراسات المبكرة بأن الاتجاه "ثنائي القطب" لجزيئات الماء يمنع تدفق البروتونات عبر آلية جروثوس. تشكك الدراسات الحديثة في هذا التفسير وتؤكد على وجود حاجز إلكتروستاتيكي كسبب لانسداد البروتون. في الرأي الأخير، فإن دوران جزيئات الماء ليس سوى أحد الآثار الجانبية للحاجز الكهروستاتيكي. في الوقت الحاضر (2008)، يعد أصل المجال الكهروستاتيكي موضع نقاش. في حين نظرت بعض الدراسات بشكل أساسي في المجال الكهربائي الناتج عن نصف حلزونات البروتين HB وHE، أكد البعض الآخر على تأثيرات الذوبان عندما يدخل البروتون في مسام الأكوابورين الضيق.
اتصال NPA
قنوات AQP، الحلزونان نصف الممتدان بواسطة أشكال NPA الملتصقة معًا حول المحور شبه المزدوج في مستوى الغشاء (الشكل 2D). يتكون تقاطع NPA هذا من جهات اتصال Vander Waals من حلقات البرولين وشبكة رابطة الهيدروجين المتشابكة التي توجه سلاسل جانبية وتطلق مجموعاتها الطرفية في المسام. في AQP1، تمركز اثنان من المياه حول تقاطع NPA، كل منهما مرتبط بالهيدروجين إلى مجموعات ND2 من NPAasparagine (Sui et al. 2001) (الشكل 2D). في AqpZ، تقوم مجموعتان من NPA asparagine ND2 بوظيفة مانح رابطة أشيدروجين لجزيء ماء مركزي، مما يحبسه في شكل يتبرع فيه هذا الماء بروابط هيدروجينية، ولكن لا يمكن قبوله من المياه المجاورة (Savage et al. 2003). InAQP0، القناة القريبة من تقاطع NPA أضيق بشكل ملحوظ، مقيدة بين زوج من مواقع انقباض التيروزين (انظر أدناه) (Harries etal. 2004). في كل من GlpF وAqpM، تقدم الوصلات NPA زوجًا من المتبرعين برابطة الهيدروجين إلى هيدروكسيل متتالي من جزيء مول الجلسرين المرتبط (الشكل 2E). ومن المثير للاهتمام، أنه تم العثور على الجلسرين فقط عند تقاطع NPA لـ GlpF، ولكن ليس AqpZ، على الرغم من تبلور كلا البروتينين في تركيزات عالية من الجلسرين. يبدو أن البيئات المحلية حول تقاطع NPA قد تشارك في هذا الاختلاف في تنسيق الماء / الجلسرين
ar/R selectivity filter
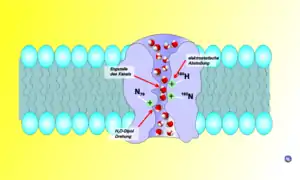
المرشح الانتقائي العطري / الأرجينين أو "ar / R" عبارة عن مجموعة من الأحماض الأمينية التي تساعد على الارتباط بجزيئات الماء واستبعاد الجزيئات الأخرى التي قد تحاول دخول المسام. إنها الآلية التي من خلالها يستطيع الأكوابورين ربط جزيئات الماء بشكل انتقائي (ومن ثم السماح لها بالمرور) ومنع الجزيئات الأخرى من الدخول. يتكون مرشح ar / R من مجموعتين من الأحماض الأمينية من الحلزون B (HB) وE (HE) ومجموعتين من الحلقة E (LE1، LE2)، من جانبي شكل NPA. موقعها المعتاد هو 8 على الجانب الخارجي من شكل NPA ؛ عادةً ما يكون الجزء الأكثر إحكامًا في القناة. يؤدي ضيقه إلى إضعاف الروابط الهيدروجينية بين جزيئات الماء، مما يمكّن الأرجينين، الذي يحمل شحنة موجبة، من التفاعل مع جزيئات الماء وتصفية البروتونات غير المرغوب فيها.
توزيع الأنواع
في الثدييات
هناك ثلاثة عشر نوعًا معروفًا من الأكوابورينات في الثدييات، وستة منها موجودة في الكلى، ولكن يُشتبه في وجود المزيد منها. تمت مقارنة الأكوابورينات الأكثر دراسة في الجدول التالي:
| Type | الموقع[31] | الوظيفة[31] |
|---|---|---|
| أكوابورين 1 |
|
إعادة إمتصاص الماء |
| أكوابورين 2 |
|
إعادة امتصاص الماء استجابة لـ ADH |
| أكوابورين 3 |
|
إعادة امتصاص الماء ونفاذية الجلسرين |
| أكوابورين 4 |
|
إعادة امتصاص الماء |
في النباتات
تؤخذ المياه في النباتات من التربة عبر الجذور، حيث تنتقل من القشرة إلى الأنسجة الوعائية. هناك ثلاثة طرق لتدفق الماء في هذه الأنسجة، والمعروفة باسم السكتات الدماغية، والممرات التناسلية وعبر الخلايا. على وجه التحديد، توجد الأكوابورينات في الغشاء الفجوي، بالإضافة إلى غشاء البلازما للنباتات ؛ يتضمن المسار العابر للخلايا نقل الماء عبر أغشية البلازما والفجوة. عندما تتعرض جذور النباتات لكلوريد الزئبق، المعروف بتثبيط الأكوابورينات، ينخفض تدفق المياه بشكل كبير بينما لا يتدفق تدفق الأيونات، مما يدعم الرأي القائل بوجود آلية لنقل المياه مستقلة عن نقل الأيونات:.
بالإضافة إلى الحفاظ على الأسمولية الطبيعية الخلوية، [مطلوب مزيد من الشرح] يمكن أن تلعب الأكوابورينات دورًا رئيسيًا في نمو الامتداد من خلال السماح بتدفق الماء إلى الخلايا المتوسعة - وهي عملية ضرورية للحفاظ على نمو النبات.
تعتبر الأكوابورينات النباتية مهمة أيضًا للتغذية المعدنية وإزالة السموم من الأيونات وكلاهما ضروري لاستتباب البورون والسيليكون والزرنيخ والبيكربونات.
تنقسم الأكوابورينات في النباتات إلى خمس فصائل فرعية متجانسة، أو مجموعات:
البروتين الداخلي لغشاء البلازما (PIP) بروتين تونوبلاست الداخلي (TIP) Nodulin-26 مثل البروتين الداخلي (NIP) البروتين الأساسي الصغير (SIP) X بروتينك الداخلي (XIP) تم لاحقًا تقسيم هذه العائلات الفرعية الخمسة إلى مجموعات فرعية تطورية أصغر بناءً على تسلسل الحمض النووي الخاص بهم. تتجمع PIPs في مجموعتين فرعيتين، PIP1 وPIP2، بينما تتجمع TIPs في 5 مجموعات فرعية، TIP1، TIP2، TIP3، TIP4 وTIP5. يتم تقسيم كل مجموعة فرعية مرة أخرى إلى أشكال إسوية على سبيل المثال PIP1 ؛ 1، PIP1 ؛ 2. نظرًا لأن تسميات الأشكال الإسوية تستند تاريخيًا إلى معايير وظيفية بدلاً من المتغيرات التطورية، فقد نشأت العديد من المقترحات الجديدة حول الأكوابورينات النباتية مع دراسة العلاقات التطورية بين الأكوابورينات المختلفة. ضمن مجموعة متنوعة من الأشكال الإسوية Aquaporin في النباتات، هناك أيضًا أنماط فريدة من التعبير الخاص بالخلية والأنسجة.
عندما يتم إسكات الأكوابورينات النباتية، تقل الموصلية الهيدروليكية والتركيب الضوئي للورقة.
عندما تحدث بوابات الأكوابورينات النباتية، فإنها توقف تدفق الماء عبر مسام البروتين. قد يحدث هذا لعدد من الأسباب، على سبيل المثال عندما يحتوي النبات على كميات قليلة من المياه الخلوية بسبب الجفاف. يتم تنفيذ بوابات الأكوابورين من خلال تفاعل بين آلية البوابات والأكوابورين، مما يؤدي إلى تغيير ثلاثي الأبعاد في البروتين بحيث يسد المسام، وبالتالي يمنع تدفق الماء عبر المسام. في النباتات، هناك نوعان على الأقل من بوابات الأكوابورين: البوابات عن طريق نزع الفسفرة لبعض بقايا السيرين، استجابة للجفاف، وبروتونات بقايا الهيستيدين المحددة، استجابة للفيضانات. تشارك الفسفرة في الأكوابورين في فتح وغلق البتلات استجابةً لدرجة الحرارة.
في الفطريات
بعض البكتيريا والعديد من الكائنات الحية الأخرى تعبر أيضًا عن الأكوابورينات.
تم اكتشاف الأكوابورينات أيضًا في الفطريات- Saccharomyces cerevisiae (الخميرة) وDictyostelium وCandida وUstilago وprotozoans- Trypanosoma وPlasmodium.
الأهمية السريرية
هناك مثالان واضحان للأمراض التي تم تحديدها على أنها ناتجة عن طفرات في الأكوابورينات:
تسبب الطفرات في جين aquaporin-2 مرض السكري الكاذب الوراثي في البشر. الفئران متماثلة اللواقح لتعطيل الطفرات في الجين aquaporin-0 تطور إعتام عدسة العين الخلقي. تم التعرف على عدد قليل من الأشخاص يعانون من نقص حاد أو كلي في Aquaporin-1. إنها صحية بشكل عام، ولكنها تظهر عيبًا في القدرة على تركيز المواد المذابة في البول والحفاظ على الماء عند حرمانها من مياه الشرب. [55] [56] الفئران التي تم حذفها من أكوابورين -1 تظهر أيضًا نقصًا في الحفاظ على المياه بسبب عدم القدرة على تركيز المواد المذابة في لب الكلى عن طريق الضرب المعاكس للتيار.
بالإضافة إلى دورها في مرض السكري الكاذب الكلوي المحدد وراثيًا، تلعب الأكوابورينات أيضًا دورًا رئيسيًا في الأشكال المكتسبة من مرض السكري الكاذب الكلوي (الاضطرابات التي تسبب زيادة إنتاج البول). يتم تنظيم Aquaporin 2 بواسطة vasopressin الذي، عندما يرتبط بمستقبل سطح الخلية، ينشط مسار إشارات cAMP. ينتج عن هذا حويصلات تحتوي على aquaporin-2 لزيادة امتصاص الماء والعودة إلى الدورة الدموية. تحور مستقبل أكوابورين 2 فازوبريسين هو سبب مرض السكري الكاذب المكتسب في الجرذان، يمكن أن يتسبب مرض السكري الكاذب الكلوي المكتسب في ضعف تنظيم الأكوابورين -2 بسبب إعطاء أملاح الليثيوم، وانخفاض تركيزات البوتاسيوم في الدم (نقص بوتاسيوم الدم) وتركيزات الكالسيوم العالية في الدم (فرط كالسيوم الدم).
تنتج تفاعلات المناعة الذاتية ضد aquaporin 4 عند البشر مرض ديفك. إذا أمكن التلاعب بالأكوابورين، يمكن أن يحل ذلك المشاكل الطبية مثل احتباس السوائل في أمراض القلب ووذمة الدماغ بعد السكتة الدماغية.
المراجع
- "The aquaporin water channels". Proc Am Thorac Soc. 3 (1): 5–13. 2006. doi:10.1513/pats.200510-109JH. PMC 2658677. PMID 16493146. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cooper G (2009). The Cell: A Molecular Approach. Washington, DC: ASM PRESS. صفحة 544. ISBN 978-0-87893-300-6. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Verkman, AS (January 2000). "Structure and function of aquaporin water channels". Am J Physiol Renal Physiol. 278 (1): F13-28. doi:10.1152/ajprenal.2000.278.1.F13. PMID 10644652. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Cooper, Geoffrey (2000). The Cell (الطبعة 2). MA: Sinauer Associates. مؤرشف من الأصل في 6 ديسمبر 2020. اطلع عليه بتاريخ 23 أبريل 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lodish, Harvey; Berk, Arnold; Zipursky, S. Lawrence (2000). Molecular Cell Biology (الطبعة 4th). New York: W. H. Freeman. ISBN 9781464183393. مؤرشف من الأصل في 6 ديسمبر 2020. اطلع عليه بتاريخ 20 مايو 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Peter Agre, 2003 Nobel Prize winner in chemistry". J. Am. Soc. Nephrol. 15 (4): 1093–5. 2004. doi:10.1097/01.ASN.0000118814.47663.7D. PMID 15034115. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The Nobel Prize in Chemistry 2003". Nobel Foundation. مؤرشف من الأصل في 18 يونيو 2018. اطلع عليه بتاريخ 23 يناير 2008. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lennon VA, Kryzer TJ, Pittock SJ, Verkman AS, Hinson SR (2005). "IgG marker of optic-spinal multiple sclerosis binds to the aquaporin-4 water channel". J. Exp. Med. 202 (4): 473–7. doi:10.1084/jem.20050304. PMC 2212860. PMID 16087714. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bichet DG (2006). "Nephrogenic diabetes insipidus" (PDF). Adv Chronic Kidney Dis. 13 (2): 96–104. doi:10.1053/j.ackd.2006.01.006. PMID 16580609. مؤرشف من الأصل (PDF) في 18 يوليو 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Aquaporin water channels: molecular mechanisms for human diseases". FEBS Lett. 555 (1): 72–8. 2003. doi:10.1016/S0014-5793(03)01083-4. PMID 14630322. S2CID 35406097. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Aquaporin-related disorders of water homeostasis". Drug News Perspect. 20 (7): 447–53. 2007. doi:10.1358/dnp.2007.20.7.1138161. PMID 17992267. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "From membrane pores to aquaporins: 50 years measuring water fluxes". J Biol Phys. 33 (5–6): 331–43. December 2007. doi:10.1007/s10867-008-9064-5. PMC 2565768. PMID 19669522. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The rate of exchange of tritiated water across the human red cell membrane". J. Gen. Physiol. 41 (2): 259–77. November 1957. doi:10.1085/jgp.41.2.259. PMC 2194835. PMID 13475690. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Goldstein DA; Solomon AK (1960-09-01). "Determination of equivalent pore radius for human red cells by osmotic pressure measurement". The Journal of General Physiology. 44: 1–17. doi:10.1085/jgp.44.1.1. PMC 2195086. PMID 13706631. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Dainty, J.; House, C. R. (1966-07-01). "An examination of the evidence for membrane pores in frog skin". The Journal of Physiology. 185 (1): 172–184. doi:10.1113/jphysiol.1966.sp007979. PMC 1395865. PMID 5965891. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The permeability to water of bimolecular lipid membranes". Journal of Theoretical Biology. 11 (3): 370–382. 1966-08-01. doi:10.1016/0022-5193(66)90099-3. PMID 5967438. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Effects of cellular acidification on ADH-induced intramembrane particle aggregates". American Journal of Physiology. Cell Physiology. 246 (1): C157–C159. 1984-01-01. doi:10.1152/ajpcell.1984.246.1.c157. ISSN 0363-6143. PMID 6320654. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Edelman, Isidore S. (25 May 1965). "Hydrogen-ion dependence of the antidiuretic action of vasopressin, oxytocin and deaminooxytocin". Biochimica et Biophysica Acta (BBA) - Biophysics Including Photosynthesis. 102 (1): 185–197. doi:10.1016/0926-6585(65)90212-8. PMID 5833400 – عبر Elsevier Science Direct. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "pH-Dependence of water and solute transport in toad urinary bladder". Kidney International. 15 (5): 513–519. 1979-05-01. doi:10.1038/ki.1979.66. ISSN 0085-2538. PMID 39188. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Zhang, RB; Logee, KA; Verkman, AS (1990-09-15). "Expression of mRNA coding for kidney and red cell water channels in Xenopus oocytes". The Journal of Biological Chemistry. 265 (26): 15375–15378. ISSN 0021-9258. PMID 2394728. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Zhang, R; Alper, S L; Thorens, B; Verkman, A S (1991-11-01). "Evidence from oocyte expression that the erythrocyte water channel is distinct from band 3 and the glucose transporter". Journal of Clinical Investigation. 88 (5): 1553–1558. doi:10.1172/JCI115466. PMC 295670. PMID 1939644. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Aquaporin CHIP: the archetypal molecular water channel". Am. J. Physiol. 265 (4 Pt 2): F463–76. 1 October 1993. doi:10.1152/ajprenal.1993.265.4.F463. PMID 7694481. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The structure of aquaporin-1 at 4.5-A resolution reveals short alpha-helices in the center of the monomer". J. Struct. Biol. 128 (1): 34–43. 1999. doi:10.1006/jsbi.1999.4177. PMID 10600556. S2CID 1076256. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The dynamics and energetics of water permeation and proton exclusion in aquaporins". Curr. Opin. Struct. Biol. 15 (2): 176–83. 2005. doi:10.1016/j.sbi.2005.02.003. hdl:11858/00-001M-0000-0012-E99D-E. PMID 15837176. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "p-(Chloromercuri)benzenesulfonate binding by membrane proteins and the inhibition of water transport in human erythrocytes". Biochemistry. 25 (7): 1535–8. 1986. doi:10.1021/bi00355a011. PMID 3011064. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The story of the discovery of aquaporins: convergent evolution of ideas--but who got there first?". Cell. Mol. Biol. (Noisy-le-grand). 52 (7): 2–5. 2006. PMID 17543213. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Benga, G. "Gheorghe Benga". Ad Astra - Online project for the Romanian Scientific Community. مؤرشف من الأصل في 25 ديسمبر 2007. اطلع عليه بتاريخ 05 أبريل 2008. الوسيط
|CitationClass=تم تجاهله (مساعدة) - A Conversation With Peter Agre: Using a Leadership Role to Put a Human Face on Science, By Claudia Dreifus, New York Times, January 26, 2009 نسخة محفوظة 6 ديسمبر 2020 على موقع واي باك مشين.
- "The structure of aquaporins". Q. Rev. Biophys. 39 (4): 361–96. 2006. doi:10.1017/S0033583506004458. PMID 17156589. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "The aquaporins". Genome Biol. 7 (2): 206. 2006. doi:10.1186/gb-2006-7-2-206. PMC 1431727. PMID 16522221. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Unless else specified in table boxes, then ref is: Walter F. Boron (2005). Medical Physiology: A Cellular And Molecular Approaoch. Elsevier/Saunders. ISBN 978-1-4160-2328-9. الوسيط
|CitationClass=تم تجاهله (مساعدة) Page 842
 صور وملفات صوتية من كومنز
صور وملفات صوتية من كومنز
 بوابة علم الأحياء
بوابة علم الأحياء
