دورة حمض الستريك
دورة حمض الستريك (بالإنجليزية: Citric acid cycle) أو حلقة حمض الليمون تعرف أيضًا بدورة الحموض ثلاثية الكربوكسيل (بالإنجليزية: TriCarboxlic acidA) أو حلقة كريبس (بالإنجليزية: Krebs Cycle) هي سلسلة من التفاعلات الكيميائية المحفزة بالأنزيمات لها أهمية أساسية في جميع الخلايا الحية التي تستخدم الأكسجين في التنفس الداخلي. تتم دورة حمض لستريك في المتقدرات في جميع خلايا الجسم.
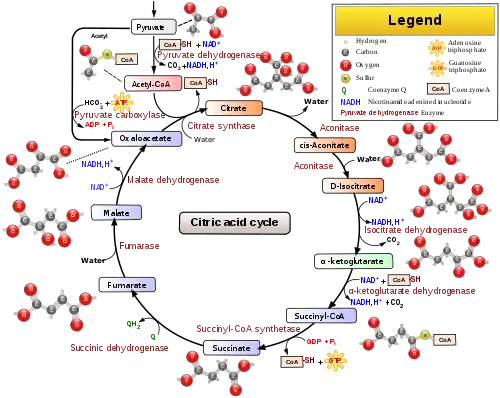
دورة حمض الستريك تسمى أيضا "دورة حمض الليمون " هي جزء من مسار الأيض الذي يحول الكربوهيدرات، و الدهون وبعض الأحماض الأمينية إلى غاز ثاني أوكسيد الكربون وماء ويولّد طاقة قابلة للاستهلاك وطاقة يختزنها في جزيئات ATP (بطارية الخلية) . دورة حمض الليمون هي المسار الثالث من أربعة مسارات استقلابية مرتبطة باستقلاب الكربوهيدرات وإنتاج ثلاثي فوسفات الأدينوسين ATP ، الثلاثة الأخرى هي أكسدة البيروفات قبله، و السلسلة التنفسية بعده. تؤمّن "دورة حمض الليمون " هياكل كربونية لبناء العديد من المركبات كبعض الأحماض الأمينية. .
الخطوات
| الجزيء | النواتج | الانزيم | نوع التفاعل | ملاحظة | |
|---|---|---|---|---|---|
| 1 | حمض أكسالوأسيتيك + أسيتيل مرافق الإنزيم-أ + H2O |
حمض الليمون + مرافق الإنزيم-أ |
سترات سينثاز | تفاعل تكاثف | rate limiting stage, extends the 4C oxaloacetate to a 6C molecule |
| 2 | حمض الليمون | cis-أكونتيز + H2O |
أكونتيز | بلمهة | reversible isomerisation |
| 3 | cis-Aconitate + H2O |
Isocitrate | تفاعل إماهة | ||
| 4 | Isocitrate + ثنائي نوكليوتيد الأدنين وأميد النيكوتين+ |
Oxalosuccinate + ثنائي نوكليوتيد الأدنين وأميد النيكوتين |
Isocitrate dehydrogenase | تفاعلات أكسدة-اختزال | generates NADH (equivalent of 2.5 ATP) |
| 5 | Oxalosuccinate | حمض ألفا كيتوغلوتارات + CO2 |
نزع كربوكسيل | irreversible stage, generates a 5C molecule | |
| 6 | حمض ألفا كيتوغلوتارات + NAD+ + CoA-SH |
Succinyl-CoA + NADH + H+ + CO2 |
α-Ketoglutarate dehydrogenase | Oxidative decarboxylation |
generates NADH (equivalent of 2.5 ATP), regenerates the 4C chain (CoA excluded) |
| 7 | Succinyl-CoA + ثنائي فوسفات الغوانوزين + فوسفات |
Succinate + CoA-SH + ثلاثي فوسفات الغوانوزين |
Succinyl-CoA synthetase | substrate level phosphorylation | or أدينوسين ثنائي الفوسفات->أدينوسين ثلاثي الفوسفات,[1] generates 1 ATP or equivalent |
| 8 | Succinate + مرافق الإنزيم Q10 (Q) |
حمض الفوماريك + ubiquinol (QH2) |
نازع هيدروجين السكسينات | أكسدة | uses ثنائي نيوكليوتيد الفلافين والأدينين as a عامل مرافق (كيمياء حيوية) (FAD->FADH2 in the first step of the reaction) in the enzyme,[2] generates the equivalent of 1.5 ATP |
| 9 | حمض الفوماريك + H2O |
L-حمض التفاح | Fumarase | تفاعل إماهة | |
| 10 | L-حمض التفاح + NAD+ |
حمض أكسالوأسيتيك + NADH + H+ | مالات ديهيدروجيناز | أكسدة | generates NADH (equivalent of 2.5 ATP) |
التفاعلات
يمكن تلخيص تفاعلات حلقة كريبس من خلال التفاعلات التالية:
التفاعل 1
تثبيت الأستيل CoA على مركب C4 لبناء مركب C6 حمض الليمون. [3] .
التفاعل 2 و3
يتم في هاذين التفاعلين عملية نزع الكربوكسيل بالإضافة إلى أكسدة ونزع H وهو ما يسمى بنزع الكربوكسيل التأكسدية.[3] .
التفاعل 4
تركيب ATP (فسفرة ADP)[3]
التفاعل 5
تفاعل أكسدة لمركب رباعي الكربون (نزع الهيدروجين)، وإرجاع ناقل +H ومرافق الإنزيم FAD←FADH2 .[3]
التفاعل 7
تفاعل أكسدة وإرجاع الناقل +NAD إلى +NADH,H .[3]
الكفاءة
إن الناتج والحاصل الأقصى النظري لـ ATP من خلال أكسدة جزيء واحد من الجلوكوز في تحلل السكر ، ودورة حمض الستريك، والفسفرة التأكسدية هو 38 (بافتراض 3 مكافئات جزيئية من ATP لكل مكافئ NADH و 2 ATP لكل FADH2). في حقيقيات النوى، يتم توليد معادلين من NADH وأربعة مكافئات من ATP في تحلل السكر، والذي يحدث في السيتوبلازم. نقل اثنين من هذه المكافئات من NADH إلى الميتوكوندريا يستهلك مكافئين من ATP، مما يقلل من صافي إنتاج ATP إلى 36. بالإضافة إلى ذلك، عدم الكفاءة في الفسفرة التأكسدية بسبب تسرب البروتونات عبر غشاء الميتوكوندريا وانزلاق ATP سينثاز / البروتون تقلل المضخة عادة من عائد ATP من NADH و FADH2 إلى أقل من العائد الأقصى النظري.[4] وبالتالي، فإن المردودات المرصودة تقترب من ~ 2.5 من ATP لكل NADH و ~ 1.5 ATP لكل FADH2 ، مما يقلل من صافي الإنتاج الإجمالي للاعبي التنس المحترفين إلى حوالي 30.[5] يوفر تقييم إجمالي عائد ATP مع نسب البروتون إلى ATP المنقحة حديثًا تقديرًا لـ 29.85 ATP لكل جزيء غلوكوز.[6]
المصادر
- Berg, JM (2002). Biochemistry - 5th Edition. WH Freeman and Company. صفحات 476. ISBN 0-7167-4684-0. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Berg et al (2002), page 501
- "تفاعلات حلقة كريبس اطلع عليه بتاريخ 12 مارس 2017 نسخة محفوظة 20 أكتوبر 2017 على موقع واي باك مشين.
- Stryer L, Berg JM, Tymoczko JL (2002). "Section 18.6: The Regulation of Cellular Respiration Is Governed Primarily by the Need for ATP". Biochemistry. San Francisco: W.H. Freeman. ISBN 0-7167-4684-0. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Porter RK, Brand MD (September 1995). "Mitochondrial proton conductance and H+/O ratio are independent of electron transport rate in isolated hepatocytes". Biochem. J. 310 (2): 379–82. doi:10.1042/bj3100379. PMC 1135905. PMID 7654171. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Rich PR (December 2003). "The molecular machinery of Keilin's respiratory chain". Biochem. Soc. Trans. 31 (Pt 6): 1095–105. doi:10.1042/BST0311095. PMID 14641005. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة أيض
بوابة أيض بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي.svg.png.webp) بوابة علم الأحياء الدقيقة
بوابة علم الأحياء الدقيقة