تطور الثدييات
هي عملية حيوية نشأت تدريجيا انفصل خلالها بين فرعي صوروبسيدا و سينابسيدا، وبدأ ذلك النفصال خلال العصر الكربوني المتأخر قبل نحو 300 مليون سنة ولا يزال هذا التطور جاريا حتى وقتنا الحالي. وتشير الأحفورات إلى أنه أثناء العصر الثلاثي الأوسط تواجدت أشباه الحيوانات الثديية. ثم ظهرت بعد ذلك الثدييات "الحقيقية " خلال العصر الثالاثي المتأخر أو ربما في أوائل العصر الجوراسي.
تطور الثدييات (بالإنجليزية: evolution of mammals)
عثر على أحفورات لحيوان سينودلفيس، وهو حيوان ثديي ذو كيس، عاش قبل 125 مليون سنة خلال العصر الطباشيري المبكر، وكان يعيش في نفس الوقت مع إيومايا الذي يعتبر أول الثدييات العليا. وبعدها بإثنين من ملايين السنين ظهر أيضا تينولوفوس أول الكظاميات وكان يعيش في المستنقعات.
بعد انقراض العصر الطباشيري-الثلاثي حيث انقرضت خلاله الديناصورات فكانت الطيور تعتبر خلائف الديناصورات. وتطورت الثدييات ذات أكياس (شبيهة الكنغر) والثدييات العليا خلال العصر الثلاثي، وتشعبت في رتب كثيرة واحتلت كل رتبة منها حيزا حيويا تستطيع العيش فيه. ومع نهاية العصر الثلاثي (الترياسي) تطورت منها جميع العائلات الثديية الحديثة.
طبقا لنظرية تطور الشعب تعتبر الثدييات هي خلائف السينابسيدات. وانفصلت السينابسيدا قبل ذلك منذ نحو 305 مليون سنة خلال العصر الكربوني من الصوروبسيدات (التي تشمل اليوم الزواحف والطيور) .[1] وأثناء العصر البرمي كانت السينابسيدات أكبر الحيوانات الفقرية على الأرض وأكثرها انتشارا.[2] الثدييات تلد والزواحف والطيور تبيض.
وخلال العصر الثلاثي أصبحت أركوصوريا -والتي كانت في السابق شعبة من الصوروبسيدات - أصبحت أكبر الفقريات على الأرض. وسادت الديناصورات التي نشأت من الأركوصورات في الأرض طوال الحقبة الوسطى ( تشمل العصور :الترياسي والجوراسي والطباشيري وتمتد من 250 مليون سنة إلى 65 مليون سنة سبقت ). وبسبب التغير البيئي وتغير المملكة النباتية خلال تلك الحقبة أضطرت الحيوانات شبيهة الثدييات إلى حياة تنشط فيها ليلا. وأدت تلك التطورات إلى نشأة حيوانات ذات صفات الثدييات مثل حرارة الجسم وظهور الشعر وزيادة حجم الدماغ . واستطاعت بعض أجناس الثدييات خلال الحقبة الوسطى المتأخرة غزو مناطق بيئية جديدة. من أمثالها "كاستروكاودا " الذي كان يعيش في المياه، و"فولاتيكوثيريوم" الذي كان شبيها بالخفاش ويطير، وكذلك "ريبينوماموس" الشبيه بالديناصورات الصغيرة .
تعتمد معلوماتنا هذه على احفورات. وكان العثور على أحفورات لثدييات وما سبقها من طوائف مباشرة من الحقبة الوسطى نادراً في الماضي، ولكن منذ التسعينيات من القرن الماضي كثر العثور على أحفوراتها وعلى الأخص في الصين.
وعن طريق تقنيات جديدة مثل تقنية التشعب الجيني الجزيئي أصبح في مقدورنا فهم بعض تطور صفات الحيوانات الثديية التي أدت إلى ظهور أنواع جديدة تعيش معنا اليوم. ومع بعض التحفطات فإن نتائج تلك التقنيات تنطبق إلى حد كبير مع ما عثر عليه من حفريات وترتيبها.
وعلى الرغم من أن الغدد الثديية من صفات الثدييات إلا أن تطور الرضاعة في الحيوان لا يزال غامضاً. وكذلك لا زلنا لا نعرف الكثير عن تطور القشرة المخية الحديثة. وتركزت أعمال الباحثين في موضوع تطور الثدييات تطور الأذن الوسطى، التي نشأت عظامها الصغيرة من عظام الفك للسلويات التي عاشت قبلها. بعض الأبحاث الأخرى تركز على تطور الأرجل والذراعين، والتطور العظمي للحنك، والفراء، والشعر و درجة حرارة الجسم.
تعريف الثدييات
يمكن التعرف على أنواع الثدييات الحالية عن طريق الإناث التي تنتج حليباً.
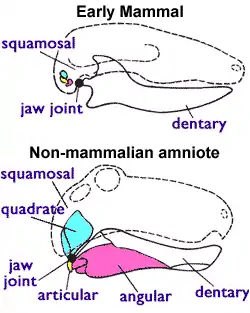
ولتصنيف أحفورة لا بد من وضع شروط أخرى حيث أن الغدد الثديية وغيرها من أعضاء الأنثى المكونة من أعضاء رخوة فهي لا توجد في الحفريات. ويستعين علماء الأحياء القديمة في التصنيف بالأذن الوسطى وهي في الثدييات الحالية وتشمل من ضمنها الثدييات ذات الكيس، ولكنها لا توجد في أشباه الثدييات (ثيرابسيدات) التي عاشت في العصر الثلاثي المبكر. توجد عظمتين للفك في السلويات وقد انزاحت عبر العصور الجيولوجية وكونت الأذن الوسطى من مطرقة وسندان فساعدت الثدييات على السمع.
كان للسلويات الأولية مفصل للفك مكون من "أرتيكولار" - وهو عظمة صغيرة في نهاية مؤخرة الفك السفلي - و عظمة "كوادراتوم" وهي عظمة صغيرة أيضا عند نهاية الفك العلوي. وجميع السلويات التي لا تنتمي إلى الثدييات مثل السحاليو التماسيح و الديناصورات وما جاء بعدهم من أحفاد، وكذلك الطيور و الثيرابسيدات، فجميعها يتميز بهذا المفصل الفكي. ولكن الثدييات تستخدم نظاما آخر للفك فهو يتكون من "دنتال" (عظمة الفك السفلي الحاملة للأسنان) وعظمة "سكواموسوم" (عظمة صغيرة من الجمجمة). وقد تحولت عظمتي كوادراتوم وأرتيكولار إلى عظمتي السندان والمطرقة في الأذن الوسطى.[3][4] وعلى عكس الفقريات الاخرى فتوجد في الثدييات بروزان أسفل الجمجمة يسمى لقمة القذالي متناسبان مع أعلى فقرة من العمود الفقري تحمل الرأس.[3]
وبغرض التعرف على الثدييات يعتمد علماء التاريخ الطبيعي على مفصل الفك بالإضافة إلى عظام الأذن الوسطى كأعضاء تميزها. وأما إضافة شروط أخرى قد توجد في أحياء مختلطة ( مثل مفصل فك من أحد أشباه الثدييات وأذن وسطى مع وجود لقمة القذالي لغير الثدييات في نفس الوقت) فهذا سوف يصعب التصنيف بلا داعي.
ويصعب أحيانا الفصل في التصنيف بين الزواحف والثدييات، عند وجود تغييرات طفيفة مرحلية قد تظهر فيما يسمى " أشكال انتقالية ".
التفرع الحيوي للثدييات
التفرع الحيوي أسفله يبين تشعبا مبسطا للفقاريات رباعية الأطراف ، ثم تناقش أسفله بعض النقاط التي يختلف عليها العلماء:
| فقاريات رباعية الأطراف (تيترابودا) |
| |||||||||||||||||||||
تغير البيئة النباتية خلال الترياسي
عندما حدث انقراض العصر البرمي الترياسي انقرضت نحو 70% من الحيوانات الفقرية التي تعيش على اليابسة وكذلك انقرضت معظم النباتات على اليابسة. وكانت النتيجة هي انهيار النظام البيئي وبالتالي الموجود الغذائي للحيوانات الباقية.[5] واستغرقت فترة استعادة الطبيعة لحيوتها نحو6 مليون سنة. وتصارعت مجموعات الحيوانات على اكتساب غذائها ونموها بين المجموعات الأخرى، كان صراعا للبقاء. ومن ضمن تلك الحيوانات كانت السينودونتات والتي يبدو أنها كانت سائدة على الأرض عند نهاية العصر البرمي.
ولكن السينودونتات خسرت معركة البقاء ضد مجموعة من نوع كان يبدو ضعيفا من صوروبسيدات وهي مجموعة أركوصورات ( وينتمي إليها التماسيح و الديناصورات و الطيور). واما ما حدث للتغير في الطبيعة النباتية التي حدثت في ذلك الوقت فهي تسمى "التغير النباتي في الترياسي". ويفكر العلماء في عدة نظريات تكون هي المستدعية لذلك التغير. وأكثر تلك النظريات احتمالا هو أن الأركوصورات التي كانت تعيش في مناطق جافة خلال العصر الثلاثي المبكر (الترياسي السفلي) كانت أكثر مقاومة لقلة وجود المياه فكانت أكثر كفاءة للمعيشة من الحيوانات الأخرى المتصارعة معها على الغذاء (جميع الصوروبسيدات لها جلد من دون مسام ويخرجون حمض البوليك بينما تخرج الثدييات وربما أيضا الثيرابسيدات تخرجان البول - ولكي يبقى حمض البوليك سائلا فهو يحتاج إلى ماء أقل مما يحتاجه البول. )[6][7] وحدث التغير النباتي الترياسي عبر فترة تعتبر طويلة . وكانت مجموعات السينودونتات من آكلات اللحوم و نوع "ليستوصوريات " كانت الغالبة في أكل اللأعشاب؛ وعندما جاء الترياسي الأوسط فكانت الأركوصورات هي السائدة واحتلت الأنماط الحياتية للحيوانات آكلة اللحوم و النباتات.
ويبدو أن التغير النباتي خلال العصر الترياسي (العصر الثلاثي) هو العامل الأكبر في تطور السينودونتات إلى الحيوانات الثديية. فما جاء من أحفاد للسينودونتات فكانت حيوانت صغيرة تصتاد غذائها ليلا وتأكل الحشرات، وبهذا استطاعت البقاء.[8] ونتيجة ذلك هو الآتي :

- كان تغير الثيرابسيدات الذي بدأ بتنوع في الأسنان وغلق جيد للفك وبسرعة، فساعده ذلك على اصتياد والإمساك بحيوانات "أرثروبودا" وكسر دروعها.
- تحتاج المعيشة الليلية عزل أحسن للحرارة وتنظيم لدرجة حرارة الجسم ، ذلك لأن أسلاف الحيوانات الثديية كانت تخرج ليلا في البرودة لاصتياد غذائها.[9]
- وكان من الضروري للبقاء على قيد الحياة التمتع بحاستي سمع وشم جيدتين.
- وكان تطور الأذن الوسطى في الثدييات مهما، ونتج عنه مفصل فكي مناسب حيث انتقلت بعض العظام المكونة للفك إلى الأذن.
- أصبح حجم المخ كبيرا حيث نمت حاستي السمع والشم بالمقارنة بنسبة حجم المخ إلى حجم الجسم. وفي نفس الوقت يحتاج المخ الأكبر إلى كمية أكبر من السعرات المتناولة مع الغذاء.[7][10] وبسبب زيادة احتياج المخ الكبير للغذاء فيستحسن اتجاه التطور تطور في العزل الحراري وتنظيم درجة حرارة الجسم، وكذلك استغلال أفضل للغذاء.
- ولكن النشاط المعيشي الليلي فله مساويء بالنسبة إلى جودة الرؤية للألوان. فانقرضت اثنتين من الأربعة أوبسنات Opsin (بروتينات لونية توجد في الخلاليا النبوتية في شبكية العين ) - فمعظم الثدييات لا يري الألوان جيدا ، ونجد ذلك في أشباه القرود مثل الليمور.[11]
تطور أهم مجموعات الثدييات
يبحث علماء التاريخ الطبيعي وعلماء تحليل الجينات الجزيئية أزمنة حدوث تغيرات وتفرعات ونوع التفرعات للحيوانات الثديية وعلى الأخص العليا منها التي بقيت حتى وقتنا الحاضر. في العادة يعتمد علماء التاريخ الطبيعي بتأريخ وجود مجموعة من الثدييات بحسب أقدم الأحفورات منها، والتي ما تحتص به من خصائص تجمع بينها. ويرى مد علماء تحليل الجينات الجزيئية أن التفرعات التطورية تكون قد انفصلت قبل أزمنة ما عثر عليه من أحفورات تخص مجموعة، وأن مجموعات كانت متشابهة في شكلها لكنها كانت تختلف بعض الشيء في جيناتها. وتمتد تلك البحوث والمناقشات أيضا إلى التعريفات ووصف كل مجموعة رئيسية عمن الثدييات العليا - ومثال عليها نجدهم يبحثون كل بطريقته في دراسة أفروثوريا محاولين التوصل إلى حل مرضي لهما.
التفرع الحيوي للثدييات العليا
يعتمد علماء التحليل الجيني الجزيئي على جينات نوعية خاصة لدراسة تفرع الكائنات وهي طريقة قريبة من الطريقة التي يتبعها علماء الحفريات والتاريخ الطبيعي والتي تعتمد على صفات معينة للأحفورات التي يعثرون عليها، فإذا تماثلت جينات كائنين أكثر من تماثلهم مع كائن حي ثالث، فيكون من اللازم إعتبار هذان الكائنان متقاربان.
التفرع التالي يبين تفرع الثدييات العليا، مع ملاحظة أنه لا يحوي ثدييات منقرضة بسبب عدم إمكانية استخلاص جينات DNS من أحفورات.
| Eutheria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
تقدير عمر تطور الثدييات العليا
تصل دراسات االتحليل الجيني إلى أن معظم رتب الحيوانات الثديية قد تفرعت قبل نحو 100 إلى 85 مليون سنة، مع تفرع العائلات الموجودة في الوقت الحاضر في زمن طويل بعد ذلك خلال الإيوسين العلوي و العصر الميوسيني السفلي.[12]
بعض علماء الحفريات يعتقدون في عدم وجود أحفورات لثدييات عليا قبل انقراض العصر الطباشيري-الثلاثي. ولكن أحفورة "ميليستس غوبينسيس" Maelestes gobiensis التي يرجع تاريخها على نحو 75 مليون سنة سبقت تنتمي إلى اليوثيريا Eutheria, ولكنها ليست لحيوان ثديي عالي.[13]
كثير من أحفورات العصر الطباشيري تبين سحالي و سمندرات و طيور و حيوانات ثديية ولكن لا يوجد بينها حيوانات ثديية عليا. ومن المحتمل أنها لم توجد في تلك الأزمنة.[14] ومع ذلك فتوجد احفورات لحيوانات عاشت قبل 85 مليون سنة من نوع الحافريات والتي من الممكن ان تكون أسلاف ذوات الحافر الحالية.[15]
ويرجع تاريخ الاحفورات الخاصة بأسلاف الحيوانات الثديية المعاصرة غالبا إلى العصر الباليوسيني المبكر (قبل 55 مليون سنة)، وقليل منها ما عاش اسلافه في العصر الطباشيري، أي قبل انقراض الديناصورات قبل 65 مليون سنة. وبعض باحثين التاريخ الطبيعي العاملين في دراسة التحليل الجينى الجزيئي يستخدم الطرق الإحصائية من أجل تتبع توالي حفريات المجموعات الحيوانية الحديثة. وقد توصلوا إلى أن الرئيسيات قد نشأت خلال العصر الطباشيري المتأخر.[16] وتؤيد الدراسات الإحصائية التي أجريت على جدول الأحفورات أن الحيوانات الثديية قد تنوعت كثيرا وزادت أحجامها سريعا خلال العصر الباليوسيني المبكر قبل 55 مليون سنة.[17][18]
تطور ملامح الثدييات
الفكوك والأذن الوسطى
تظهر مستحاثة Hadrocodium، والتي تعود إلى العصر الجوراسي المبكر، أول دليل واضح على مفاصل الفك الكامل للثدييات والأذنين الوسطى لها، حيث يتكون مفصل الفك من عظام الفك السفلي والعظم القشري، بينما يوجد العظم المربعي والمفصلي في الأذن الوسطى، ويُعرفان باسم عظمي المطرقة والسندان.
توضح إحدى تحليلات Teinolophos من فصيلة الكظاميات أن لهذا الحيوان مفصل فك ما قبل الثدييات، أي يتكون من العظم المربعي والعظم الزاوي، كما يوضح التحليل أن الأذن الوسطى للثدييات النهائية تطورت مرتين بشكل مستقل، في فصيلتي الكظاميات والوحشيات، لكن هذه الفكرة كانت موضع خلاف.[19]
شارك اثنان من مؤلفي هذا الاقتراح في تأليف ورقة لاحقة أعادت تفسير نفس السمات كدليل على أن حيوان الـ Teinolophos كان خلد ماء مكتمل النمو، مما يعني أنه ربما امتلك مفصلاً للفك والأذن الوسطى.[20]
الإرضاع
اتُفق أن وظيفة الإرضاع الأصلية هي الحفاظ على رطوبة البيض. وتستند هذه الفكرة في الأساس على فصيلة الكظاميات (الثدييات التي تضع البيض).[21][22][23] من المعروف أن البيض السلوي قادرٌ على التطور بعيداً عن الماء. لكن معظم بيوض الزواحف تحتاج إلى الرطوبة كي لا تجف.
لا تحتوي الكظاميات على حلمات، لكنها تفرز الحليب من بقعة شعر على بطونها.
تُغطى بيوض الكظاميات أثناء الحضانة بمادة لزجة مجهولة الأصل. وتتألف قشرتها من ثلاث طبقات قبل وضع البيوض. ثم تظهر طبقة رابعة بتركيبة مختلفة عن تركيبة الطبقات الثلاث الأصلية بعد وضع البيوض. وتُنتج الطبقة الرابعة والمادة اللزجة من الغدد الثديية.
وإن كان الأمر كذلك، فربما نحصل على تفسيرٍ لسبب إفراز الكظاميات للحليب من بقعٍ شعرية. فمن الأسهل نشر الرطوبة ومواد أخرى على البويضة من منطقة واسعة تملك شعراً أكثر من الحلمة الصغيرة العارية.
أظهرت الأبحاث اللاحقة ظهور الكازين عند سلف الثدييات المشترك منذ نحو 200 إلى 310 مليون عام.[24] يدفعنا ذلك إلى التساؤل ما إذا كان إفراز مادة ما للحفاظ على رطوبة البيضة يشير إلى حدوث الإرضاع عند الثيرابسيدات. هناك ثدييات صغيرة تدعى Sinocodon –والتي تُعد مجموعة شقيقة لجميع الثدييات اللاحقة –لها أسنان أمامية حتى عند أصغر أفرادها، بالإضافة إلى الفك المتحجر بشكل سيء. ربما لم تقم هذه الحيوانات بعملية الإرضاع[25]، أي ربما تطورت الرضاعة مباشرة أثناء الانتقال من مرحلة ما قبل الثدييات إلى مرحلة الثدييات. ومع ذلك، تُظهر الـ Tritylodontidae –وهو حيوان قاعدي –أدلة واضحة على حدوث عملية الإرضاع.[26] كما يُظهر الـ Morganucodonta –وهو حيوان قاعدي وثديي الشكل –دليلاً على حدوث عملية الإرضاع.[27]
الشعر والفراء
أول دليل واضح على الشعر أو الفراء هو مستحاثات الـ Castorocauda والـ Megaconus، منذ 164 مليون سنة في منتصف العصر الجوراسي.[28] نظراً لامتلاك كلا الحيوانين طبقةً مزدوجة من الشعر وشعر وطلية سفلية، فمن المفترض أن يملك آخر سلفٍ مشترك لهما الميزات الشعرية ذاتها. يجب أن يكون هذا الحيوان من العصر الترياسي لأنه كان سلفاً للـ Tikitherium من العصر الترياسي أيضاً.[28]
في منتصف الخمسينيات، فسّر بعض العلماء الثقب (أو الممرات) في الفك العلوي وقادمة الفك العلوي (وهي عظام صغيرة أمام الفك العلوي) لدى كلبيات الأسنان كقنوات تزود الشعرات الأنفية عن الحيوان (أو الشوارب) بالأوعية الدموية والأعصاب، واقترحوا أن هذا دليلٌ على وجود الشعر أو الفراء.[29][30]
ووضحوا أيضاً أن الثقوب لا تدل بالضرورة على امتلاك الحيوان للشعيرات الأنفية. فالسحلية الحديثة Tupinambis لديها ثقوب تكاد تكون مماثلة لتلك الموجودة لدى كلبيات الأسنان غير الثديية كالـ Thrinaxodon.[31][32] ومع ذلك، تواصل المصادر الشائعة إسناد الشوارب إلى الـ Thrinaxodon.[33]
اعتُبرت الحفريات الأثرية، عن طريق الخطأ، والعائدة للعصر الترياسي السفلي أنها بصمة تظهر الشعر لدى كلبيات الأسنان [34]، لكن هذا الادعاء دُحض واعتبر باطلاً.[35]
في دراسة للفتحات القحفية وارتباط الأعصاب الوجهية بالشوارب لدى الثدييات البعيدة، وُجد أن الـ Prozostrodontia –وهو من الأسلاف الصغيرة والمباشرة للثديات –تملك شوارب (شعيرات) شبيهة بتلك الموجودة لدى الثدييات، ولكن الثيرابسيدات الأقل تطوراً لا تملك شعيرات متحركة، وقد لا تملك شعيرات على الإطلاق.[36] ربما تطور الفراء من الشعيرات[37]، أما الشعيرات فربما تطورت كرد فعل على نمط الحياة الليلية أو/و الاختباء في الكهوف.
الأطراف المنتصبة
لم نستطع فهم عملية تطور الأعضاء المنتصبة عند الثدييات بشكل كامل، حيث تمتلك الكظاميات الحالية والمتحجرة على سبيل المثال أعضاء متمددة. ويعتقد العلماء أن وضعية الطرف غير المتمدد محدودة لدى الإسفينيات الشمالية، وهي مجموعة تحتوي الوحشيات باستثناء اللانابيات. ويتفق العلماء أن خاصية الوقوف ظهرت لدى الـ Sinodelphys والـ Eomaia، أي أن وضعية الوقوف هذه ظهرت منذ 125 مليون سنة في العصر الطباشيري المبكر. ويعتقد العلماء أن الثديات الأولى امتلكت أطرافاً أمامية منتصبة على عكس الأطراف الخلفية الأكثر تمدداً. ولاحظوا استمرار هذا الاتجاه لدى المشيميات والجرابيات الحديثة.[38]
ذوات الدم الحار
إن الدم الحار مصطلح معقد ومبهم، فهو يتضمن بعض أو جميع الأنواع التالية: داخلية الحرارة: وهي القدرة على توليد الحرارة داخلياً بدلاً من ممارسة سلوكيات معينة كالتشمس أو ممارسة النشاط العضلي.
ثابتة الحرارة: وتعني ثبات درجة حرارة الجسم. حيث تملك معظم الأنزيمات مجالاً مثالياً لدرجات الحرارة كي تعمل. وتنخفض الفعالية بسرعة خارج هذا المجال. يحتاج الكائن ثابت الحرارة إلى امتلاك أنزيمات تعمل بشكل جيد في مجال صغيرٍ من درجات الحرارة.
ذوات الدم الحار: تتميز هذه الكائنات بمعدل أيضٍ (استقلاب) مرتفع، وتحديداً في حالة الراحة. ويتطلب ذلك درجة حرارة مرتفعة نسبياً، وإمكانية الحفاظ على درجة الحرارة هذه. وسبب ذلك هو معامل Q10: أي تنخفض سرعة العمليات الكيميائية الحيوية إلى النصف تقريباً إن انخفضت درجة حرارة الحيوان بمقدار 10 درجات مئوية. لا يعلم العلماء كثيراً عن الآليات الداخلية لتلك الحيوانات المنقرضة. وتركز معظم النقاشات على ثبات الحرارة وذوات الدم الحار. لكنهم اتفقوا أن داخلية الحرارة قد تطورت في البداية عند مندمجات الأقواس كالديساينودونت، حيث امتلكت تلك الحيوانات جسداً متناسقاً ساعدها في احتجاز الحرارة[39]، بالإضافة إلى عظام مليئة بالأوعية الدموية وقنوات هافرس.[40] وربما امتلكت شعراً أيضاً.[41] وهناك اقتراحٌ ظهر في الآونة الأخيرة يوضح أن داخلية الحرارة قد تطورت منذ ظهور أفعواني العيون.[42]
تملك الكظاميات الحديثة درجة حرارة داخلية منخفضة مقارنة مع الثدييات المشيمية والجرابية. حيث تبلغ نحو 32 درجة مئوية (أو 90 فهرنهايت).[43] ووفقاً لتطور السلالات القوسي، لم تكن درجة حرارة هذه الثدييات أقل من تلك التي امتلكتها الكظاميات. هناك دليلٌ وفقاً لعلم الأحياء الخلوي يُظهر أن معدل الاستقلاب المنخفض لدى الكظاميات ليس سوى صفة ثانوية متطورة.[44]
المراجع
- "Amniota - Palaeos". مؤرشف من الأصل في 03 مارس 2016. اطلع عليه بتاريخ 27 ديسمبر 2010. الوسيط
|CitationClass=تم تجاهله (مساعدة); تحقق من التاريخ في:|تاريخ أرشيف=(مساعدة) - "Synapsida overview - Palaeos". مؤرشف من الأصل في 27 يناير 2010. اطلع عليه بتاريخ 27 ديسمبر 2010. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Mammalia: Overview - Palaeos نسخة محفوظة 27 يونيو 2017 على موقع واي باك مشين.
- Cowen, R. (2000). History of Life. Oxford: Blackwell Science. صفحة 432. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Olenekian Age of the Triassic - Palaeos". مؤرشف من الأصل في 30 ديسمبر 2017. اطلع عليه بتاريخ 28 أغسطس 2013. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Kermack, D.M.; Kermack, K.A. (1984). The evolution of mammalian characters. Croom Helm. ISBN 0709915349. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Brain power". New Scientist. 2006. مؤرشف من الأصل في 28 أغسطس 2017. اطلع عليه بتاريخ 27 ديسمبر 2010. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Cynodontia Overview – Palaeos". مؤرشف من الأصل في 17 يناير 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ruben, J.A., and Jones, T.D. (2000). "Selective Factors Associated with the Origin of Fur and Feathers". American Zoologist. 40 (4): 585–596. doi:10.1093/icb/40.4.585. مؤرشف من الأصل في 29 أغسطس 2009. الوسيط
|CitationClass=تم تجاهله (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Raichle, M.E.; Gusnard, D.A. (2002-08-06). "Appraising the brain's energy budget". PNAS. 99 (16): 10237–10239. doi:10.1073/pnas.172399499. مؤرشف من الأصل في 25 يوليو 2008. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Travis, J (2003). "Visionary research: scientists delve into the evolution of color vision in primates". Science News. 164 (15). مؤرشف من الأصل في 08 يوليو 2012. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bininda-Emonds, O.R.P.; Cardillo, M.; Jones, K.E.; et al. (2007). "The delayed rise of present-day mammals". Nature. 446 (446): 507–511. doi:10.1038/nature05634. مؤرشف من الأصل في 14 يناير 2012. الوسيط
|CitationClass=تم تجاهله (مساعدة); Explicit use of et al. in:|الأخير4=(مساعدة) - Dinosaur Extinction Spurred Rise of Modern Mammals نسخة محفوظة 01 نوفمبر 2017 على موقع واي باك مشين.
- Benton, M.J. (1999). "Early origins of modern birds and mammals: molecules vs. morphology". BioEssays. 21: 1043–1051. doi:10.1002/(SICI)1521-1878(199912)22:1<1043::AID-BIES8>3.0.CO;2-B. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Archibald, J.D. (1996). "Fossil Evidence for a Late Cretaceous Origin of "Hoofed" Mammals". Science. 272 (5265): 1150–1153. doi:10.1126/science.272.5265.1150. مؤرشف من الأصل في 26 يناير 2005. اطلع عليه بتاريخ 08 سبتمبر 2008. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Martin, R.D.; Soligo, C.; Tavaré, S. (2007). "Primate Origins: Implications of a Cretaceous Ancestry" (PDF). Folia Primatologica. 78 (78): 277–296. doi:10.1159/000105145. مؤرشف من الأصل (PDF) في 10 أبريل 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) — Ein ähnliches Papier derselben Autoren ist kostenlos im Netz verfügbar: New light on the dates of primate origins and divergence نسخة محفوظة 26 فبراير 2009 على موقع واي باك مشين. - Alroy J. (1999). "The fossil record of North American mammals: evidence for a Paleocene evolutionary radiation". Systematic biology. 48 (1): 107–18. doi:10.1080/106351599260472. PMID 12078635. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Archibald J.D., and Deutschman D.H. (2001). "Quantitative Analysis of the Timing of the Origin and Diversification of Extant Placental Orders". Journal of Mammalian Evolution. 8 (2): 107–124. doi:10.1023/A:1011317930838. مؤرشف من الأصل في 14 أكتوبر 2012. اطلع عليه بتاريخ 24 سبتمبر 2008. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Rich, T.H.; Hopson, J.A.; Musser, A.M.; Flannery., T.G.F. & Vickers-Rich, P. (11 February 2005). "Independent Origins of Middle Ear Bones in Monotremes and Therians". Science. 307 (5711): 910–914. Bibcode:2005Sci...307..910R. doi:10.1126/science.1105717. PMID 15705848. الوسيط
|CitationClass=تم تجاهله (مساعدة) For other opinions see "Technical comments" linked from same Web page - Rowe, T.; Rich, T.H.; Vickers-Rich, P.; Springer, M. & Woodburne, M.O. (January 2008). "The oldest platypus and its bearing on divergence timing of the platypus and echidna clades". Proceedings of the National Academy of Sciences. 105 (4): 1238–1242. Bibcode:2008PNAS..105.1238R. doi:10.1073/pnas.0706385105. PMC 2234122. PMID 18216270. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Oftedal, O.T. (2002). "The mammary gland and its origin during synapsid evolution" (PDF). Journal of Mammary Gland Biology and Neoplasia. 7 (3): 225–252. doi:10.1023/A:1022896515287. PMID 12751889. مؤرشف من الأصل (PDF) في 21 سبتمبر 2013. الوسيط
|CitationClass=تم تجاهله (مساعدة) -
Oftedal, O.T. (2002). "The origin of lactation as a water source for parchment-shelled eggs" (PDF). Journal of Mammary Gland Biology and Neoplasia. 7 (3): 253–266. doi:10.1023/A:1022848632125. PMID 12751890. مؤرشف من الأصل في 5 يونيو 2012. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lactating on Eggs نسخة محفوظة April 14, 2009, على موقع واي باك مشين.
- Borders, R.; Robertson, W. (1993). "Washington's pesticide panel: process and product". Veterinary and Human Toxicology. 35 (3): 258–259. PMID 8351802. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Mammals of the Mesozoic: The least mammal-like mammals نسخة محفوظة 10 نوفمبر 2012 على موقع واي باك مشين.
- Hu, Yaoming; Meng, Jin; Clark, James M (2009). "A New Tritylodontid from the Upper Jurassic of Xinjiang, China". Acta Palaeontologica Polonica. 54 (3): 385–391. doi:10.4202/app.2008.0053. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Elsa Panciroli; Roger B. J. Benson; Stig Walsh (2017). "The dentary of Wareolestes rex (Megazostrodontidae): a new specimen from Scotland and implications for morganucodontan tooth replacement". Papers in Palaeontology. in press. doi:10.1002/spp2.1079.
- P Bajdek, Microbiota and food residues including possible evidence of pre‐mammalian hair in Upper Permian coprolites from Russia, G. 2015, doi:10.1111/let.12156
- Brink, A.S. (1955). "A study on the skeleton of Diademodon". Palaeontologia Africana. 3: 3–39. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Kemp, T.S. (1982). Mammal-like reptiles and the origin of mammals. London: Academic Press. صفحة 363. ISBN 978-0-12-404120-2. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bennett, A.F.; Ruben, J.A. (1986). "The metabolic and thermoregulatory status of therapsids". In Hotton III, N; MacLean, P.D.; Roth, J.J.; et al. (المحررون). The ecology and biology of mammal-like reptiles. Washington: Smithsonian Institution Press, Washington. صفحات 207–218. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Estes, R. (1961). "Cranial anatomy of the cynodont reptile Thrinaxodon liorhinus". Bulletin of the Museum of Comparative Zoology. 125 (1253): 165–180. مؤرشف من الأصل في 12 فبراير 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Thrinaxodon: The Emerging Mammal". National Geographic Daily News. February 11, 2009. مؤرشف من الأصل في 7 نوفمبر 2017. اطلع عليه بتاريخ 26 أغسطس 2012. الوسيط
|CitationClass=تم تجاهله (مساعدة); غير مسموح بالترميز المائل أو الغامق في:|ناشر=(مساعدة) - Ellenberger, Paul (1976). "Une piste avec traces de soies épaisses dans le Trias inférieur a moyen de Lodève (Hérault, France): Cynodontipus polythrix nov. gen., nov. sp. les Cynodontes en France". Geobios. 9 (6): 769–787. doi:10.1016/S0016-6995(76)80078-2. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Olsen, Paul E. (2012): Cynodontipus: A procolophonid burrow – not a hairy cynodont track (Middle-Late Triassic: Europe, Morocco, Eastern North America). Program for Northeastern Section – 47th Annual Meeting (18–20 March 2012) Hartford, Connecticut, the Geological society of America نسخة محفوظة 09 مارس 2016 على موقع واي باك مشين. [وصلة مكسورة]
- Benoit, J.; Manger, P. R.; Rubidge, B. S. (9 May 2016). "Palaeoneurological clues to the evolution of defining mammalian soft tissue traits". Scientific Reports. 6: 25604. Bibcode:2016NatSR...625604B. doi:10.1038/srep25604. PMC 4860582. PMID 27157809. مؤرشف من الأصل في 10 أبريل 2020. اطلع عليه بتاريخ 14 سبتمبر 2016. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Switek, Brian. "Getting to the Root of Fur". National Geographic. مؤرشف من الأصل في 28 أغسطس 2017. اطلع عليه بتاريخ 24 نوفمبر 2014. الوسيط
|CitationClass=تم تجاهله (مساعدة) -
Kielan−Jaworowska, Z.; Hurum, J.H. (2006). "Limb posture in early mammals: Sprawling or parasagittal" (PDF). Acta Palaeontologica Polonica. 51 (3): 10237–10239. مؤرشف من الأصل (PDF) في 8 يونيو 2019. اطلع عليه بتاريخ 24 سبتمبر 2008. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bakker 1975
- JENNIFER BOTHA-BRINK and KENNETH D. ANGIELCZYK, Do extraordinarily high growth rates in Permo-Triassic dicynodonts (Therapsida, Anomodontia) explain their success before and after the end-Permian extinction?, Version of Record online: 26 JUL 2010 doi:10.1111/j.1096-3642.2009.00601.x
- Microbiota and food residues including possible evidence of pre-mammalian hair in Upper Permian coprolites from Russia Piotr Bajdek1, Martin Qvarnström2, Krzysztof Owocki3, Tomasz Sulej3, Andrey G. Sennikov4,5, Valeriy K. Golubev4,5 andGrzegorz Niedźwiedzki2 Article first published online: 25 NOV 2015 doi:10.1111/let.12156
- "Ancestry of mammalian 'warm-bloodedness' revealed". www.sciencedaily.com. Society of Vertebrate Paleontology. October 29, 2015. مؤرشف من الأصل في 3 أبريل 2019. اطلع عليه بتاريخ 29 أكتوبر 2015. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Paul, G.S. (1988). Predatory Dinosaurs of the World. New York: Simon and Schuster. صفحة 464. ISBN 978-0-671-61946-6. مؤرشف من الأصل في 15 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - J.M. Watson; J.A.M. Graves (1988). "Monotreme Cell-Cycles and the Evolution of Homeothermy". Australian Journal of Zoology. 36 (5): 573–584. doi:10.1071/ZO9880573. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة ثدييات
بوابة ثدييات بوابة علم الأحياء التطوري
بوابة علم الأحياء التطوري

{kind=link}