استدامة المرحلة اليرقية
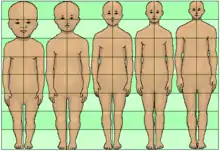
احتفاظ البالغين بالصفات التي توجد فقط في الأطفال شيء مهم في البيولوجيا التطورية، وبالتالي في التطور. ويحدث ذلك بطريقتين. في استدامة المرحلة اليرقية[1][2][3][4] (التي تسمى أيضا : الطفولية/الحداثية)[5] يتباطأ أو يتأخر التطور الفسيولوجي (أو الجسدي) للكائن الحي (عادة حيوان) . كما تم العثور على استدامة المرحلة اليرقية في البشر المعاصرين.[6]
في المقابل، في التكاثر المبكر، يحدث التطور الجنسي بشكل أسرع.[7] تؤدي كلتا العمليات إلى التشكل العضوي للجنين، كنوع من التفاوت الزمني.[8] بعض الكُتاب تعرّف التشكل العضوي بالإبقاء على الصفات اليرقية، كما نرى في السلمندر.[9][10][11]
التاريخ والتأصيل
تنحدر أصول مفهوم استدامة المرحلة اليرقية وصولا إلى الكتاب المقدس (كما يناقش أشلي مونتاجو) والشاعر ويليام وردزورث "الطفل هو والد الرجل" (كما يناقش باري بوجين). تم استخدام المصطلح نفسه في عام 1885 من قبل يوليوس كولمان في وصفه لنضج قنفذ البحر في حين تبقى في مرحلة المائية مشابهة للخلايا الشرغوفية كاملة مع الخياشيم، على عكس البرمائيات البالغين الآخرين مثل الضفادع والعلاجيم.[12]
تم استعار كلمة استدامة المرحلة اليرقية من "Neotenie" الألمانية[؟]، وهذه الأخيرة التي أطلقت من قبل كولمان من "νέος" اليونانية (الأجسام القريبة من الأرض، "الطفل") وτείνειν (teínein "،تعني التمديد").[13] لعكس كلمة "مستدام المرحلة اليرقية"، تستخدم السلطات المختلفة "تنضج الخصائص الجنسية" [14][15][16] ، يشير بوجين إلى أن كولمان كان ينوي للمعنى أن يكون "الحفاظ على الشباب"، ولكن الخلط الواضح ان teínein اليونانية مع تينيري اللاتينية، التي كان لها المعنى الذي أراده "الاحتفاظ"، لهذا فإن الكلمة جديدة تعني "الاحتفاظ بالشباب (حتى سن البلوغ) ".[12]
في عام 1926، وصف لويس بولك استدامة المرحلة اليرقية بأنها عملية كبرى في تكوين الإنسان.[12] وأشار ستيفن جاي غولد في كتابه 1977 "كتاب تطور الجنين والتأريخ العرقي" [17] أن حسابات بولك شكلت محاولة "علمية" لتبرير العنصرية والتمييز على أساس الجنس، ولكنه اعترف أن بولك كان معه الحق في الفكرة الأساسية أن البشر تختلف من الرئيسيات الأخرى في أنهم ينضجون جنسيا في المرحلة الطفولية من نمو[؟] الجسم.[12]
في البشر

استدامة المرحلة اليرقية في البشر هو تباطؤ أو تأخير نمو الجسم، مقارنة مع الرئيسيات غير البشرية، مما يؤدى إلى بعض السمات، مثل الرأس الكبير، والوجه المسطح، وقصر الذراعين والساقين نسبيا. قد تكون هذه التغيرات المستدامة اليرقية ناتجة عن الانتقاء الجنسي في التطور البشري. في المقابل، قد تكون سمحت لتنمية القدرات البشرية مثل التواصل العاطفي. ومع ذلك، يمتلك البشر أيضا أنوف كبيرة نسبيا وسيقان طويلة، كصفات نضج شكلي (لا استدامة يرقية). وقد اقترح بعض منظرين التطورية أن استدامة المرحلة اليرقية كانت سمة رئيسية في التطور البشري. يعتقد ستيفن جاي غولد أن "القصة التطورية" للبشر هي واحدة حيث كنا "نحتفظ إلى مرحلة البلوغ بملامح الطفولية (الإحداث) في الأصل من أجدادنا".[18] يعكس جي.بي.إس هالدين فرضية غولد بالقول «اتجاه تطوري كبير في البشر" هو" إطالة أكبر للطفولة والتأخر في النضج." [5] وقال دلبرت دي. تيسين أن "استدامة المرحلة اليرقية يصبح أكثر وضوحا كما تطورت القرود في وقت مبكر إلى أشكال لاحقة" وأن هذه قرود "تتطور في اتجاه وجه مسطح."[19] ومع ذلك، في ضوء بعض الجماعات التي تستغل الحجج القائمة حول استدامة المرحلة اليرقية لدعم العنصرية، كما قال غولد "أن المشروع بأكمله لترتيب المجموعات حسب درجة استدامة المرحلة اليرقية غير مبرر في الأساس" [20](غولد، 1996، ص. 150). قال دوغ جونز هذا الاتجاه في التطور البشري نحو استدامة المرحلة اليرقية قد يكون ناجما عن طريق الانتقاء الجنسي في التطور البشري لصفات الوجه في النساء من قبل الرجال الذين يعانون من استدامة المرحلة اليرقية مما أدى إلى وجوه الرجال بأنه "منتج قبلي" للانتقاء الجنسي عن وجوه الإناث .[21]
في الحيوانات المستأنسة
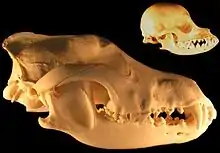
يمكننا رؤية استدامة المرحلة اليرقية في الحيوانات المستأنسة مثل الكلاب والفئران.[22] هذا لأن هناك المزيد من الموارد المتاحة، وأقل منافسة لتلك الموارد، ومع انخفاض المنافسة تنفق الحيوانات طاقة أقل للحصول على هذه الموارد. مما يعطيها الفرصة حتى تنضج وتتكاثر بسرعة أكبر من نظرائها البرية.[22] البيئة التي تربي الحيوانات المستأنسة تحدد ما إذا كان أو لم تكن استدامة المرحلة اليرقية موجودة في تلك الحيوانات. استدامة المرحلة اليرقية التطورية يمكن أن تنشأ في فصيلة[؟] ما عندما تحدث تلك الظروف، وتصبح الأنواع ناضجة جنسيا قبل موعد "النمو الطبيعي". تفسير آخر لاستدامة المرحلة اليرقية في الحيوانات المستأنسة يمكن أن يكون انتخاب بعض الخصائص السلوكية. يرتبط السلوك بعلم الوراثة الذي بالتالي يعني أنه عندما يتم اختيار سمة سلوكية، يمكن اختيار سمة جسدية أيضا بسبب آليات مثل اختلال توازن الربط. في كثير من الأحيان، يتم تحديد السلوكيات من أجل استئناس بسهولة أكبر لنوع ما؛ العدوانية في بعض الأنواع تأتي مع سن البلوغ عندما يكون هناك حاجة للتنافس على الموارد. إذا لم يكن هناك حاجة للمنافسة، إذن ليست هناك حاجة للعدوان. يمكن اختيار الخصائص السلوكية الحداثية أن تؤدي إلى استدامة المرحلة اليرقية في الخصائص الجسمانية لأنه، على سبيل المثال، مع قلة الحاجة إلى سلوكيات مثل العدوان ليست هناك حاجة لصفات متقدمة من شأنها أن تساعد في هذا المجال. الصفات التي يمكن أن تصبح مستدامة المرحلة اليرقية بسبب نقص العدوانية قد يكون الخطم أقصر والحجم العام أصغر بين الأفراد المستأنسة. بعض الصفات المشتركة المستدامة المرحلة اليرقية المادية في الحيوانات المستأنسة (وخصوصا الكلاب والخنازير والقوارض والقطط، وحتى الثعالب) ما يلي: الآذان المرنة، والتغيرات في الدورة التناسلية، وذيول مجعدة، تلوين ذو بقع، فقرات[؟] أقل أو أقصر، وعيون كبيرة، وجبهته مدورة، آذان كبيرة، وخطم قصير.[23][24]
عندما توسع دور الكلاب من مجرد كونها الكلاب العاملة إلى أن تصبح مصاحبة، بدأ البشر أيضا تربية الكلاب بناء على انتقائية استدامة المرحلة اليرقية الشكلية، وهذه التربية الانتقائية ل "استدامة المرحلة اليرقية أو الاحتفاظ بالخصائص الحداثية" كان له تأثير في تعزيز العلاقة بين البشر والكلاب. [25] البشر ربت الكلاب للحصول على مزيد من "الصفات الجسدية الحداثية" عند البلوغ مثل الخطوم قصيرة والعيون الواسعة التي توجد في الجراء، لأن الناس عادة ما تعتبر هذه الصفات أكثر جاذبية. بعض سلالات الكلاب مع الخطوم القصيرة ورؤوس كبيرة مثل كوموندور، وسانت برنارد وماريما كلب الراعي أكثر استدامة للمرحلة اليرقية من حيث الشكل من السلالات الأخرى من الكلاب.[26] يعد " فارس الملك تشارلز السبنيلي" مثال على الاختيار لاستدامة المرحلة اليرقية، لأنها تملك العيون الكبيرة والأذنين على شكل قلادة والقدمين المدمجة، مما يعطيها التشكل المماثل بين الجراء والبالغين. [25]
في عام 2004، خلصت الدراسة التي قامت باستخدام 310 من جماجم الذئاب وأكثر من 700 جماجم كلاب تمثل 100 سلالة، أن تطور جماجم الكلاب عموما لا يمكن وصفها من خلال عمليات التفاوت الزمني مثل استدامة المرحلة اليرقية على الرغم من أن بعض سلالات الكلاب تحتفظ بالخصائص الطفولية/ الحداثية التي تشبه جماجم الذئاب الإحداثية.[27] بحلول عام 2011، تم الاكتشاف من قبل الباحث نفسه ببساطة "الكلاب ليست ذئاب محتفظة بخصائص حداثية".[28]
في الأنواع الأخرى

قد لوحظت استدامة المرحلة اليرقية في العديد من الأنواع الأخرى. ومن المهم أن نلاحظ الفرق بين استدامة المرحلة اليرقية الجزئية والكاملة عند النظر في الأنواع الأخرى من أجل التمييز بين الصفات الإحداثية التي تقتصر فائدتها فقط في المدى القصير، والصفات التي توفر المنافع طوال حياة الكائن الحي.[29][30] هذا قد يقدم بعض التبصر حول سبب استدامة المرحلة اليرقية في تلك الأنواع. استدامة المرحلة اليرقية الجزئية هي الإبقاء على الشكل اليرقي بعد السن المعتاد للنضج مع إمكانية تطوير أجهزة التكاثر الجنسي مبكرا، ولكن في نهاية المطاف فإن الكائن الحي لا يزال ينضج في اتجاه البلوغ. هذا ما يمكن ملاحظته في الضفدع الأخضر. ويمكن أن نرى استدامة المرحلة اليرقية الكاملة في "عفريت الماء" وبعض أنواع "السمنلدر المخطط"، التي لا تبقى في شكلها اليرقي طوال مدة حياتهم. يسلك الضفدع الأخضر استدامة المرحلة اليرقية جزئيا عندما يؤخر عملية نضجه خلال فصل الشتاء لأنه ليس من المفيد بالنسبة له التحور إلى الشكل البالغ إلا إذا كان هناك المزيد من الموارد المتاحة حيث يمكن العثور على تلك الموارد بسهولة أكبر في شكل يرقات. هذا من شأنه أن تندرج تحت كل من الأسباب الرئيسية للاستدامة المرحلة اليرقية. الطاقة اللازمة من أجل البقاء في فصل الشتاء تكون مكلفة جدا بالنسبة للبالغين، لذلك يسلك الكائن خصائص يرقية حتى الوقت الذي تصبح فيه قادرة على البقاء على قيد الحياة أفضل ككائن بالغ. يحتفظ السلمندر المخطط بخصائصه اليرقية لسبب مماثل، لكن هذا الاحتفاظ دائم بسبب نقص الموارد المتاحة طوال فترة حياته. وهذا مثال آخر لسبب بيئي للاستدامة المرحلة اليرقية في الأنواع التي لازالت محتفظة بخصائصها الطفولية لأن البيئة تحد من قدرة الكائن الحي أن يأتي بالكامل في شكل بالغ . تظهر بعض الأنواع القليلة من الطيور استدامة جزئية للمرحلة اليرقية . هناك أمثلة على هذه الأنواع هي طائر ماناكن (شيروزيفيا الخطي ) والماناكن الأزرق (شيروزيفيا كوداتا). تحتفظ الذكور من النوعين بالريش الطفولي في مرحلة البلوغ، ولكنها تفقده في نهاية المطاف مرة واحدة عندما تكون ناضجة تماما.[31] في بعض أنواع الطيور النادرة غالبا ما يرتبط الإبقاء على ريش الطفولي بأوقات طرح الريش في كل الأنواع. من أجل ضمان عدم وجود تداخل بين طرح الريش والتزاوج بعض الأحيان، قد تظهر الطيور استدامة المرحلة اليرقية جزئية في ما يخص ريش الذكور لكي لا يحصل الذكور على ريش البالغين المشرق قبل أن تكون الإناث على استعداد للتزاوج. في هذه الحالة، فإن استدامة المرحلة اليرقية موجودة لأنه ليست هناك حاجة للذكور إلى تساقط الريش في وقت مبكر، وأنها ستكون مضيعة للطاقة في محاولة التزاوج في حين أن الإناث لا تزال غير ناضجة.
يمكننا رؤية استدامة المرحلة اليرقية عادة في الحشرات غير الطائرة مثل الإناث في نظام ملتويات الأجنحة. وقد تطورت هذه الصفة "عدم الطيران" في هذه الحشرات عبر عدة مراحل منفصلة. البيئات التي قد ساهمت في تطور منفصل من هذه الصفة هي: المرتفعات العالية، العزلة في الجزر، والحشرات التي تعيش في المناخ البارد [32].قد تكون هذه العوامل البيئية مسئولة عن سمة عدم الطيران، لأنه في هذه الحالات يكون من غير الملائم لهذه الحشرات أن تتشتت، لذلك يُفضل عدم الطيران بسبب الحدود التي تؤدى للتشتت. أيضا، في درجات الحرارة المنخفضة تفقد الحرارة بسرعة أكبر من خلال أجنحة[؟]، وبالتالي فإن الظروف تفضل انعدام الطيران. نقطتين أخريين من النقاط الرئيسية للملاحظة حول الحشرات هي أن الإناث في مجموعات معينة تصبح ناضجة جنسيا دون التحول الشكلي إلى مرحلة البلوغ، وبعض الحشرات التي تنمو في ظروف معينة لا تتكون لديها الأجنحة. يرتبط انعدام الطيران في بعض الحشرات الإناث بارتفاع الخصوبة[؟]، وهذا من شأنه زيادة اللياقة البدنية للفرد لأن الأنثى تنتج ذراري أكثر، وبالتالي تنقل لها الكثير من الجينات.[32] في تلك الحالات، تحدث استدامة المرحلة اليرقية لأنه أكثر فائدة للإناث بانعدام الطيران، من أجل الحفاظ على الطاقة وبالتالي زيادة الخصوبة لديها. يعتبر المن مثال هام على الحشرات التي قد لا تتطور لديها الأجنحة إطلاقا بسبب وضع البيئة الخاصة بهم. إذا كانت الموارد وفيرة فليست هناك حاجة لنمو الأجنحة والانتشار. عندما تكون التغذية في النبات المضيف وفيرة، قد لا تنمو الأجنحة[؟]، وتبقى المن على النبات العائل طوال فترة حياتها؛ ومع ذلك، إذا أصبحت الموارد قليلة، فإن ذريتها قد تتطور لديها الأجنحة من أجل الانتشار على نباتات مضيفة أخرى.[33]
تشترك بيئتين في الميل نحو استدامة المرحلة اليرقية، هي: الارتفاعات العالية، والبيئات الباردة لأن الأفراد مستدامي المرحلة اليرقية تكون لديهم لياقة بدنية أعلى من الأفراد التي تتحور إلى الشكل البالغ. وذلك لأن الطاقة اللازمة للتحول مكلفة جدا على حساب لياقة الفرد، أيضا تفضل الظروف استدامة المرحلة اليرقية بسبب قدرة الأفراد على الاستفادة من الموارد المتاحة بسهولة أكبر.[34] يمكن رؤية هذا الاتجاه في المقارنة بين أنواع السمندل على الارتفاعات المنخفضة والمرتفعة. الأفراد مستدامي المرحلة اليرقية يكون ليدهم قدرة أكبر للبقاء على قيد الحياة وكذلك خصوبة أعلى من السلمندر الذي كان قد تحول إلى النموذج البالغ في الارتفاعات العالية والبيئة الأكثر برودة.[34] الحشرات في البيئات الأكثر برودة تميل إلى إظهار استدامة المرحلة اليرقية في الطيران لأن الأجنحة لديها مساحة واسعة وتفقد الحرارة بسرعة، وبالتالي فإنه ليس من المفيد للحشرات في تلك البيئة أن تتحور إلى الشكل البالغ.[32]
العديد من أنواع السمندل، والبرمائيات بشكل عام، ومن المعروف أن لها خصائص استدامة المرحلة اليريقة بسبب البيئة التي يعيشون فيها. وقنفذ البحر هو نوع من السمندل التي تحتفظ بالشكل المائي الطفولي في جميع مراحل البلوغ، وهو مثال ممتاز على الاستدامة الكاملة للمرحلة اليرقية . الخياشيم هي السمة اليرقية المشتركة في البرمائيات التي يتم الاحتفاظ بها بعد النضج. وكمثال على هذا نرى المقارنة بين السمندل النمر والنيوت خشن البشرة، وكلاهما يحتفظ الخياشيم في مرحلة البلوغ.[29]
الشمبانزي القزم (البابون) تشترك في العديد من الخصائص الجسمانية مع البشر. ومن أبرز الأمثلة وجماجمه المستدامة اليرقية، [35] حيث أن شكل الجمجمة عندهم لا يتغير في مرحلة البلوغ. لكنه يزيد فقط في الحجم. ويرجع ذلك إلى ازدواج الشكل الجنسي والتغير التطوري في توقيت نضجها.[35] يصبح اليافعون ناضجين جنسيا قبل أن تتطور أجسادهم بشكل كامل في مرحلة البلوغ، ونظرا لبعض المميزات الانتقائية بقي الهيكل اليرقي للجمجمة في الأجيال اللاحقة.
في بعض الأنواع، تكاليف الطاقة تؤدي إلى استدامة المرحلة اليرقية، كما هو الحال في أسر الحشرات : جيريدي، ديلفاسيديا، والخنفساء الأرضية، العديد من الأنواع في هذه الأسر لديها أجنحة يرقية صغيرة، أو بلا أجنحة على الإطلاق.[33] وبالمثل، تبدل بعض أنواع الكريكيت أجنحتها في مرحلة البلوغ، [36] بينما في خنافس أوزوبيمون، الذكور (يعتقد أنها أول مثال على استدامة المرحلة اليرقية في الخنافس) هي أصغر بكثير من الإناث، من خلال التوالد الداخلي.[37] بينما في النمل الأبيض "كالوترمس فلافيكوليس"، نرى استدامة المرحلة اليرقية في الإناث خلال طرح الأجنحة[؟].[38]
في الأنواع الأخرى، الظروف البيئية تسبب استدامة المرحلة اليرقية، كما هو الحال في السمندل الشمالية الغربية(السلمندر الجراسيلية)، حيث يرتبط وجودها على ارتفاعات عالية بزيادة الميول نحو الاستدامة اليرقية، ربما للمساعدة في الحفاظ على الطاقة على النحو المذكور أعلاه.[39] وبالمثل، تم العثور على استدامة المرحلة اليرقية في عدد قليل من الأنواع من عائلة القشريات اشنوميزديا، والتي تعيش في مياه المحيطات العميقة.[40]
انظر أيضا
المصادر
- Neoteny. (2012). Dictionary.com. Retrieved September 19, 2012, from link نسخة محفوظة 04 مارس 2016 على موقع واي باك مشين.
- Neoteny. (2012). Your dictionary. Retrieved September 19, 2012, from link. Pronunciation is shown as (nēŏtˈnē) and in IPA notation it is /niːˈɒtni/ as can be inferred from the Pronunciation Key. نسخة محفوظة 30 أبريل 2017 على موقع واي باك مشين.
- Neoteny. (2012). Oxford Dictionaries. Retrieved September 19, 2012, from link. Pronunciation is shown as /nēˈätn-ē/ and was changed to IPA notation using ويكيبيديا:أصد للإنجليزية and the vowels of fleece, lot, and happy. نسخة محفوظة 08 أغسطس 2016 على موقع واي باك مشين.
- Neoteny. (2012). Merriam-Webster. Retrieved September 19, 2012, from link. Pronunciation is shown as ē.ˈä.tə.nē\ and was changed to IPA notation using the pronunciation guide and ويكيبيديا:أصد للإنجليزية. نسخة محفوظة 06 يناير 2017 على موقع واي باك مشين.
- Montagu, A. (1989). Growing Young. Bergin & Garvey: CT.
- Being More Infantile May Have Led to Bigger Brains نسخة محفوظة 08 أغسطس 2017 على موقع واي باك مشين.
- Volkenstein, M. V. 1994. Physical Approaches to Biological Evolution. Springer-Verlag: Berlin, . نسخة محفوظة 16 ديسمبر 2019 على موقع واي باك مشين.
- Ridley, Mark (1985). Evolution. Blackwell. مؤرشف من الأصل في 26 مايو 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Whiteman, H.H. (1994). "Evolution of facultative paedomorphosis". Quarterly Review of Biolology. 69: 205–221. doi:10.1086/418540. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Schell, S. C. Handbook of Trematodes of North America North of Mexico, 1985, pg. 22
- Ginetsinskaya, T.A. Trematodes, Their Life Cycles, Biology and Evolution. Leningrad, USSR: Nauka 1968. Translated in 1988, . نسخة محفوظة 16 ديسمبر 2019 على موقع واي باك مشين.
- Bogin, Barry (1999). Patterns of Human Growth. Cambridge University Press. صفحات 157–169. ISBN 978-0-521-56438-0. مؤرشف من الأصل في 25 يناير 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Neoteny, The Free Dictionary. 2011. Accessed April 30, 2011. نسخة محفوظة 22 سبتمبر 2018 على موقع واي باك مشين.
- Henke, W. (2007). Handbook of paleoanthropology, Volume 1. Springer Books, NY.
- Hetherington, R. (2010). The Climate Connection: Climate Change and Modern Human Evolution. Cambridge University Press.
- Hall, B.K., Hallgrímsson, B. Monroe, W.S. (2008). Strickberger's evolution: the integration of genes, organisms and populations. Jones and Bartlett Publishers: Canada.
- Gould, Stephen Jay (1977). Ontogeny and Phylogeny. Cambridge, Massachusetts: Belknap (Harvard University Press). ISBN 0-674-63940-5. مؤرشف من الأصل في 16 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Gould, S.J. A Biological Homage to Mickey Mouse.
- Thiessen, D.D. (1997). Bittersweet destiny: the stormy evolution of human behavior. Transaction Publishers, N.J.
- Gould, S.J. (1996). The mismeasure of man. W.W. Norton and Company, N.Y.
- Jones, D.; et al. (1995). "Sexual selection, physical attractiveness, and facial neoteny: Cross-cultural evidence and implications [and comments and reply]". Current Anthropology. 36 (5): 723–748. doi:10.1086/204427. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Price, E. (1999). "Behavioral development in animals undergoing domestication". Applied Animal Behaviour Science. 65 (3): 245–271. doi:10.1016/S0168-1591(99)00087-8. مؤرشف من الأصل في 24 سبتمبر 2015. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bertone, J. (2006). Equine geriatric medicine and surgery. Saunders, MI.
- Trut, L. N. (1999). "Early canid domestication: the farm-fox experiment". American Scientist. 87 (2): 160–169. doi:10.1511/1999.2.160. الوسيط
|CitationClass=تم تجاهله (مساعدة) - McGreevy, P.D. & Nicholas, F.W. (1999). Some Practical Solutions to Welfare Problems in Dog Breeding. In Animal Welfare. 8: 329-341.
- Beck, A.M. & Katcher, A.H. (1996). Between Pets and People: The Importance of Companionship. West Lafayette, Indiana: Purdue University Press. ISBN 1-55753-077-7
- Drake, Abby Grace, "Evolution and development of the skull morphology of canids: An investigation of morphological integration and heterochrony" (January 1, 2004). Doctoral Dissertations Available from Proquest. Paper AAI3136721. link نسخة محفوظة 29 ديسمبر 2017 على موقع واي باك مشين.
- Drake, Abby Grace (2011). "Dispelling dog dogma: An investigation of heterochrony in dogs using 3D geometric morphometric analysis of skull shape". Evolution & Development. 13 (2): 204. doi:10.1111/j.1525-142X.2011.00470.x. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Swingle, W. (1922). "Experiments on the metamorphosis of neotenous amphibians". Journal of Experimental Zoology. 36 (4): 397–421. doi:10.1002/jez.1400360402. الوسيط
|CitationClass=تم تجاهله (مساعدة) - AmphibiaWeb - Ambystoma tigrinum نسخة محفوظة 05 فبراير 2017 على موقع واي باك مشين.
- Foster, M. (1987). "Delayed maturation, neoteny, and social system differences in two manakins of genus Chiroxyphia". Evolution. 41 (3): 547–558. doi:10.2307/2409256. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Barbosa, P.; et al. (1989). "Life-history traits of forest-inhabiting flightless Lepidoptera". American Midland Naturalist. 122 (2): 262–274. doi:10.2307/2425912. JSTOR 2425912. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Harrison, R (1980). "Dispersal polymorphisms in insects". Annual Review of Ecology and Systematics. 11: 95–118. doi:10.1146/annurev.es.11.110180.000523. JSTOR 2096904. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Snyder, R (1956). "Comparative Features of the Life Histories of Ambystoma gracile (Baird) from Populations at Low and High Altitudes". Copeia. 1956 (1): 41–50. doi:10.2307/1439242. JSTOR 1439242. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Shea, B. T. (1983). "Paedomorphosis and Neoteny in the Pygmy Chimpanzee". Science. 222 (4623): 521–522. doi:10.1126/science.6623093. JSTOR 1691380. PMID 6623093. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Harrison, R (1980). "Dispersal Polymorphisms in Insects". Annual Review of Ecology and Systematics. 11: 95–118. doi:10.1146/annurev.es.11.110180.000523. JSTOR 2096904. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Jordal, B. H.; Beaver, R. A.; Normark, B. B.; Farrell, B. D. (2002). "Extraordinary sex ratios and the evolution of male neoteny in sib-mating Ozopemon beetles". Biological Journal of the Linnean Society. 75: 353–360. doi:10.1046/j.1095-8312.2002.00025.x. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Soltani-Mazouni, N.; Bordereau, C. (1987). "Changes in the cuticle, ovaries and colleterial glands during the pseudergate and neotenic molt in Kalotermes flavicollis (FABR.) (Isoptera : Kalotermitidae)". International Journal of Insect Morphology and Embryology. 16 (3–4): 221–225. doi:10.1016/0020-7322(87)90022-5. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Eagleson, G.; McKeown, B. (1978). "Changes in thyroid activity of Ambystoma gracile (Baird) during different larval, transforming, and postmetamorphic phases". Canadian Journal of Zoology. 56 (6): 1377–1381. doi:10.1139/z/78-190. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Brokeland, W.; Brandt, A. (2004). "Two new species of Ischnomesidae (Crustacea: Isopoda) from the Southern Ocean displaying neoteny". Deep-Sea Research Part II. 51 (14–16): 1769–1785. doi:10.1016/j.dsr2.2004.06.034. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة زواحف وبرمائيات
بوابة زواحف وبرمائيات بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الحيوان
بوابة علم الحيوان
