أوليغومايسين
الأوليغومايسينات (بالإنجليزية: Oligomycins) هي ماكروليدات تنشأ من البكتيريا المتسلسلة ويمكن أن تكون سامة لكائنات حية أخرى.
| أوليغومايسين A | |
|---|---|
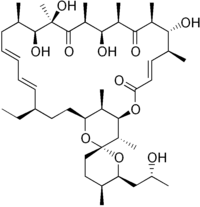 أوليغومايسين | |
| الاسم النظامي (IUPAC) | |
(1R,4E,5'S,6S,6'S,7R,8S,10R,11R,12S,14R,15S,16R,18E,20E,22R,25S,27R,28S,29R)-22-ethyl-7,11,14,15-tetrahydroxy-6'-[(2R)-2-hydroxypropyl]-5',6,8,10,12,14,16,28,29-nonamethyl-3',4',5',6'-tetrahydro-3H,9H,13H-spiro[2,26-dioxabicyclo[23.3.1]nonacosa-4,18,20-triene-27,2'-pyran]-3,9,13-trione | |
| المعرفات | |
| رقم CAS | 1404-19-9 |
| بوب كيم (PubChem) | 6450197 |
مواصفات الإدخال النصي المبسط للجزيئات
| |
| |
| الخواص | |
| الكتلة المولية | 791.062 غ/مول. |
| المخاطر | |
| صحيفة بيانات سلامة المادة | MSDS at Fermentek |
| في حال عدم ورود غير ذلك فإن البيانات الواردة أعلاه معطاة بالحالة القياسية (عند 25 °س و 100 كيلوباسكال) | |
الوظيفة
يستخدم كمضاد حيوي.
اوليغومايسن A عبارة عن مثبط لإنزيم تصنيع الطاقة. في أبحاث الفسفتة الاكسيدية، يستخدم لإيقاف المرحلة الثالثة للتنفس الخلوي. يقوم اوليغومايسن A بتثبيط إنزيم تصنيع الطاقة عن طريق سد قناته البروتونية التي تعمل على تحويل ال ADP →ATP .
إن تثبيط تصنيع الطاقة من قبل اوليغومايسن A يؤدي إلى خفض سريان الإلكترونات عبر سلسلة نقل الإلكترون، لكن لا يتوقف سريان الإلكترونات تماما بسبب عملية تسمى ب تسرب البروتونات أو عن طريق فك ربط المايتوكندريا.[1] هذه العملية تحدث نتيجة الانتشار الميسر للبروتونات إلى قالب المايتوكندريا بمساعدة بروتين فك ربط مثل الثرموجينين.
قد يؤدي تعاطي اوليغومايسن إلى نسب عالية من اللاكتات متراكمة في الدم والبول.
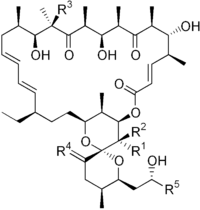 | |||||
|---|---|---|---|---|---|
| R1 | R2 | R3 | R4 | R5 | |
| Oligomycin A | CH3 | H | OH | H,H | CH3 |
| Oligomycin B | CH3 | H | OH | O | CH3 |
| Oligomycin C | CH3 | H | H | H,H | CH3 |
| Oligomycin D (Rutamycin A) |
H | H | OH | H,H | CH3 |
| Oligomycin E | CH3 | OH | OH | O | CH3 |
| Oligomycin F | CH3 | H | OH | H,H | CH2CH3 |
| Rutamycin B | H | H | H | H,H | CH3 |
| 44-Homooligomycin A | CH2CH3 | H | OH | H,H | CH3 |
| 44-Homooligomycin B | CH2CH3 | H | OH | O | CH3 |
موقع ارتباط - الاوليجومايسن
اوليجومايسن معترف به كمثبط فاعل لانزيم الATP سينثاز في المتكندريا منذ عام 1958 عندما تم إفادته من قبل هينري لاردي (henry lardy et al) .[3]
في ستينات القرن العشرين تمت درسات في مختبر افرايم راكر (EfraimRacker) أظهرت أن أنزيم الATP سينثاز في المتوكندريا يمكن تقسيمه إلى جزئين f1
(coupling factor 1/عامل اقتران 1 )
الذي يحتوي موقع التحفيز لتصنيع ال ATP ، وf0
( coupling factor 0/ عامل اقتران 0 )
الجزء الذي يكون حساساً للاوليجومايسن [4] [5]، ولكن بالرغم من 50 سنة من الدراسات على f1f0 للATP سنثازفي المتكندريا ، لم تستطع فهم حقيقة ارتباط الاوليجومايسن بالجزء f0 . لذلك تم اقتراح ، وجود موقع ارتباط للاليجومايسن على الوحدة الفرعية (c (subunit-c على الجزء f0 من ال ATP سينثاز ، هذه الوحدة الفرعية هي عبارة عن بروتين غشائي أساسي ، تتكون من حِلْزين 1و 2 ، تعبران الغشاء الداخلي للمتكندريا . الوحدة الفرعية c ، والتي يتم تمثل حلقات متشابهة تتكون من 10 وحدات فرعية في ال ATP سينثاز في الخميرة وثمانية وحدات فرعية في ال ATP سينثاز في البقر .[6][7] حلقة ال(c (c-ring تعتبر مكون أساسي في محرك التربين للبروتاونات في ATP سينثاز ، والمقترن في حركة ال البروتونات مع تدرج الجهد . وقد افتُرض أن الكربوكسيل الأساسي ل Glu59 في الحِلِزُّ 2 من الوحدة الفرعية c في الخميرة يشارك مباشرة في حركة البروتونات من السايتوسول إلى مَطْرِسُ المتكندريا (mitochondrial matrix) خلال تصنيع ال ATP . توجد السلسلة الكاربوكسيل الجانبية لل Glu59 في منتصف الِحلز 2 ، مما يضعها في طبقة الليبيد الثنائية على شكل بروتوني مغلق . [8]
بينما افتُرض ان الوحدة الفرعية (a (subunit-a تشكل اثنان من القنوات النصفية الذائبة (aqueous half-channles) ، والتي تسمح للبروتونات بالوصول إلى كربوكسيل ال Glu59 في شكل السلسلة المفتوح ، والذي بدوره يسمح بعمليات إضافة وإزالة البروتونات (protonation and deprotonation) .
و على هذه الأساس تم اقتراح أنه في الATP سنثاز السليم ، يرتبط الاوليجومايسن بالحلقة c الواقعة في قنوات البروتونات ، والذي بدوره يقوم بإيقاف الحركة الإنتقالية للبروتونات ، عن طريق منع الوصول للكربوكسيل الأساسي . وتم اقتراح أيضاً أن موقع الارتباط المؤطر في الاوليجومايسن ،هو موقع ارتباط لأي دواء مثبط يرتبط في الحلقة c للبكتيريا ، والمثبطات التي ترتبط في v0 (مضاهي لf0 ) لانزيم ال ATPase . بسبب سعة الانتشار لهذه المثبطات . ومن المحتمل أن يمثل موقع الارتباط للاوليجومايسن ، قاعدة أساسية لتطور الأدوية في المستقبل [9]
المراجع
- Jastroch M, Divakaruni AS, Mookerjee S, Treberg JR, Brand MD (2010). "Mitochondrial proton and electron leaks". Essays in biochemistry (47): 53–67. doi:10.1042/bse0470053. PMID 20533900. الوسيط
|CitationClass=تم تجاهله (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Nakata, Masaya; Ishiyama, Takashi; Akamatsu, Shinichi; Hirose, Youichi; Maruoka, Hiroshi; Suzuki, Rika; Tatsuta, Kuniaki (1995). "Synthetic studies on oligomycins. Synthesis of the oligomycin B spiroketal and polypropionate portions". Bulletin of the Chemical Society of Japan. 68 (3): 967–89. doi:10.1246/bcsj.68.967. الوسيط
|CitationClass=تم تجاهله (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - 3. Lardy HA, Johnson D, McMurray WC (1958) Antibiotics as tools for metabolic studies.I. A survey of toxic antibiotics in respiratory, phosphorylative and glycolytic systems.ArchBiochemBiophys 78:587–597.
- 4. Racker E (1963) A mitochondrial factor conferring oligomycin sensitivity on solublemitochondrial ATPase. BiochemBiophys Res Commun 10:435–439.
- 6. Kagawa Y, Racker E (1966) Partial resolution of the enzymes catalyzing oxidativephosphorylation. 8. Properties of a factor conferring oligomycinsensitivity on mitochondrialadenosine triphosphatase. J BiolChem 241:2461–2466.
- 7. Watt IN, Montgomery MG, Runswick MJ, Leslie AG, Walker JE (2010) Bioenergetic costof making an adenosine triphosphate molecule in animal mitochondria. Proc NatlAcadSci USA 107:16823–16827.
- 8. Stock D, Leslie AGW, Walker JE (1999) Molecular architecture of the rotary motor inATP synthase. Science 286:1700–1705.
- 9. Pogoryelov D, et al. (2010) Microscopic rotary mechanism of ion translocation in theF(o) complex of ATP synthases. Nat ChemBiol 6:891–899.
- 10. Symersky J, Osowski D, Walters DE, Mueller DM.Proc Natl AcadSci U S A. 2012 Aug 28;109(35):13961-5. doi: 10.1073/pnas.1207912109. Epub 2012 Aug 6.
 بوابة الكيمياء
بوابة الكيمياء بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة صيدلة
بوابة صيدلة بوابة طب
بوابة طب
