مركب إكسوسوم
مركب إكسوسوم[1] (مـ1) هو مركب متعدد البروتين داخل خلايا معقدة قادر على إتلاف أنواع مختلفة من جزيئات RNA (حمض نووي ريبوزي). وقد عُثِرَ على مركب الإكسوسوم في كل من الخلايا حقيقيات النوى والعَتائِق، في حين أن في البكتيريا مركب أبسط يسمى ديغرادوسوم ينفذ وظائف مماثلة.
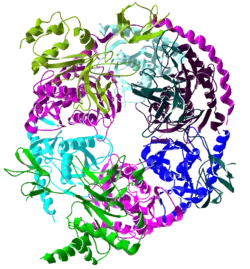
تحتوي نواة الإكسوسوم على بنية حلقية سداسية الأضلاع ترتبط بها بروتينات أخرى. فَفي الخلايا حقيقيات النوى، يوجد مركب الإكسوسوم في سيتوبلازم، ونواة الخلية، وعلى وجه الخصوص في النويّات، على الرغم من أن البروتينات المختلفة تتفاعل مع مركب الإكسوسوم في هذه المقصورات التي تؤدي إلى إتلاف الحمض النووي الريبوزي وتنظم نشاط المركب للركيزات المخصصة لهذه المقصورات الخلوية. وتَشْمَلُ ركائز الإكسوسوم الرنا رسول، والرنا الريبوسومي، والعديد من أنواع الرنا الصغيرة. الإكسوسوم لديه وظيفة إكستريبونوكليوليتيكية (exoribonucleolytic)، وهذا يعني أنه يَحُطُّ من الحمض النووي الريبوزي بدءا من نهاية واحدة (في هذه الحالة النهاية 3')، وفي حقيقيات النوى أيضا وظيفته إندوريبونوكليوليتيكية (endoribonucleolytic)، وهذا يعني أن الحمض النووي الريبوزي ينفصم في مَواقِع داخل الجُزَيْء.
العديد من البروتينات في الإكسوسوم هي هدف الأجسام الذاتية المضادة عند المرضى الذين يعانون من أنواع محددة من أمراض المناعة الذاتية (وخاصة التهاب العضل التصلبي) وبعض العلاج الكيميائي بمضاد المئيضة لمرض السرطان عن طريق منع نشاط الإكسوسوم. وبالإضافة إلى ذلك، فإن الطفرات في عنصر الإكسوسوم 3 هي سبب نَقْصُ التَّنَسُّج الجِسْرِيٌّ المُخَيخِي ومرض الخلايا العصبية الحركية الشوكية.
الاكتشاف
اكْتُشِفَ الإكسوسوم لأول مرة باعتباره ريبونوكلياز في عام 1997 في فطريات الخميرة، وهو غالبا ما يستخدم كنموذج حي.[2] ولم يمض وقت طويل، بعد ذلك في عام 1999، أُدْرِكَ أن الإكسوسوم كان في الواقع هو ما يعادل فطريات الخميرة لمركب وُصَِف بالفعل في الخلايا البشرية يسمى PM/Scl complex، والذي حُدِّدَ كمستضد ذاتي عند المرضى الذين يعانون من أمراض المناعة الذاتية في السنوات السابقة.[3] سمحت تَنْقِية "PM/Scl complex" بالتعرف على المزيد من بروتينات الإكسوسوم عند الإنسان، وفي نهاية المطاف وُصِفَتْ جميع مكونات المُركب.[4][5] في عام 2001، سَمَحَت الكمية المتزايدة من بيانات الجينوم التي أصبحت متاحة بالتنبؤ بِبروتينات الإكسوسوم في العَتائِق، على الرغم من أن الأمر استغرق سنتين سابقتين قبل أن يُنَقى مُركب الإكسوسوم لأول مرة من كائن حي.[6][7]
بنية
بروتينات أساسية
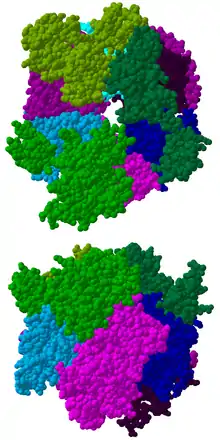
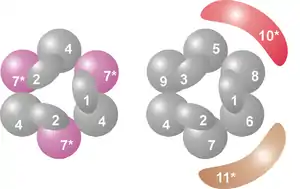
لَدَى المركب جوهرياً بنية حلقية تتكون من ستة بروتينات تنتمي جميعها إلى نفس فئة الريبونوكلياز (RNase)، مثل بروتين RNase PH.[8] فَفي العَتائِق هناك نوعان من بروتينات-PH المختلفة (تسمى Rrp41 وRrp42)، كل منهما يَخْضَعُ لترتيب في ثلاث مرات بالتناوب، حيث أن مركبات الإكسوسوم اليوكاريوتيكية لديها ستة بروتينات مختلفة والتي بدورها تشكل بنية حلقية،[9][10] من هذه البروتينات حقيقيات النوى الستة، ثلاثة تشبه بروتين العَتائِق Rrp41 والبروتينات الثلاثة الأخرى هي أكثر مماثلة لبروتين العَتائِق Rrp42.[11]
تقع على رأس هذه البنية الحلقية ثلاثة بروتينات رابطة للرنا لديها مجال S1 (غير محدد)، بالإضافة إلى ذلك هناك بروتينات نطاق تناظر K.[8] وفي حقيقيات النوى، هناك ثلاثة بروتينات مختلفة من متلازمة "S1" في بنية حلقية، في حين في العَتائِق إما بروتين أو بروتينين من مختلف بروتينات "S1" يمكن أن تكون جزءا من الإكسوسوم (على الرغم من أن هناك دائمًا ثلاث وحدات فرعية S1 تتعلق بالمركب).[12]
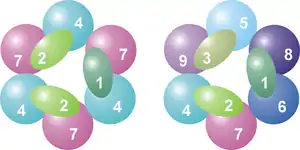
هذه البنية عبارة عن بنية حلقية مشابهة تماماً لتلك التي تتكون منها بروتينات RNase PH و PNPase. في البكتيريا، وبروتين RNase، الذي يشارك في معالجة tRNA، مما يشكل بِنْيَةً حَلقية هيكساميريكية تتكون من ستة بروتينات متطابقة من RNase PH.[13] وفي حالة PNPase، وهو بروتين الحمض النووي الفوسفوري الموجود في البكتيريا والبلاستيدات الخضراء والميتوكندريون عند بعض الكائنات حقيقيات النوى، هناك مجالين من RNase، كل من مجال متلازمة S1 و KH RNA وهما جزءين من بروتين واحد، والذي يشكل مركب ثلاثي له بنية مماثلة تقريبا لتلك البنية عند الإكسوسوم.[14] وبسبب هذا التشابه الكبير في كل من المجالات البروتينية والبنيوية، يعتقد أن هذه المركبات يمكن أن تكون ذات صلة تطورية ولها سلف مشترك.[15] في البكتيريا، يوجود البروتين RNase PH منفصل حيث يشارك في معالجة الحمض النووي الريبوزي الناقل، الذي ثُبِّتَ على بنية مماثلة لبنية ستة أعضاء حلقية، ولكن في هذه الحالة تتكون من 6 وحدات فرعية من البروتينات المتطابقة.[16] بروتينات الإكسوسوم مثل RNase PH، و PNPase و RNase PH تنتمي كلها إلى عائلة RNases PH من RNases وإكسوريبونوكلياز فسفوروليتي، وهذا يعني أنها تستخدم الفوسفات غير العضوي لإزالة النيوكليوتيدات من النهاية 3' لجزيئات الحمض النووي الريبوزي.[8]
بروتينات مقترنة
بالإضافة إلى هذه البروتينات الأساسية التسعة لمركب الإكسوسوم، فإن بروتينان آخرين يرتبطان غالباً بالمركب في الكائنات حقيقيات النوى، واحد منهما هو Rrp44، وهو ريبونوكلياز محلول مائي، الذي ينتمي إلى عائلة RNase R من إكسوريبونوكلياز المتحلل بالماء (نوكلياز يستخدم المياه لفَسخ الروابط النوكليوتيدية).
بالإضافة إلى كونه إنزيم فريد من نوع الأكسدة، فإن Rrp44 لديه أيضًا نشاط مغاير للخلايا العصبية، والذي يتواجد في مجال منفصل عن البروتين.[17][18] وفي الخميرة، يرتبط Rrp44 بكافة مركبات الإكسوسوم وله دور حاسم في نشاط مركب الإكسوسوم في الخميرة [19]
في حين وجود نديد الإنسان من البروتين، لم يُعثَر على أدلة لفترة طويلة أن نديد الإنسان يرتبط مع مركب الإكسوسوم عند الإنسان.[8] ومع ذلك، في عام 2010، اكْتُشِفَ أن البشر لديهم ثلاثة نديدات من Rrp44 ويمكن ربط اثنين منها مع مركب الإكسوسوم، وعلى الأرجح أن هذين البروتينين يمكن أن يتحللان لركائز رنا مختلفة بسبب توطينه الخلوي المختلف، حيث يتم تحديد موضع واحد في السيتوبلازم (Dis3L1) والآخر في النواة (Dis3).[20][21]
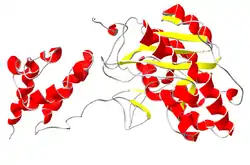
يسمى البروتين الثاني المشترك المرتبط Rrp6 (في الخميرة) أو PM / Scl-100 (في الإنسان)، مثل Rrp44، هذا البروتين هو عبارة عن نبتة تحويلية مائية، ولكن في هذه الحالة هو من عائلة بروتين RNaseD.[22] إن بروتين PM/Scl-100 هو في الغالب جزء من مركبات الإكسوسوم في نواة الخلايا، ولكن يمكن أن يشكل جزءًا من مركب الإكسوسوم السيتوبلازمي أيضًا.[23]
بروتينات تنظيمية
وبغض النظر عن هذين الجزئين الفرعيين المرتبطين بإحكام، تتفاعل العديد من البروتينات مع مركب الإكسوسوم في كل من السيتوبلازم ونواة الخلايا، هذه البروتينات المرتبطة فضفاضة قد تنظم نشاط وخصوصية مركب الإكسوسوم، في السيتوبلازم، يتفاعل الإكسوسوم مع بروتينات عناصر AU-الغنيّة (مثل KRSP وTTP)، التي يمكنها تعزيز أو منع إتلاف mRNAs. يرتبط الجسيم النووي ببروتينات الارتباط RNA (مثل MPP6/Mpp6 وC1D/Rrp47 عند البشر/الخميرة) المطلوبة لمعالجة ركائز معينة.[8]
بالإضافة إلى البروتينات المفردة، تتفاعل مركبات البروتينات الأخرى مع الإسوسوم، واحد منها هو سيتوبلازميك مركب سكي، والذي يتضمن رنا هيليكاز (سكي 2) ويشارك في إتلاف الحمض النووي الريبوزي الناقل.[24] في النواة، تتم معالجة rRNA و snoRNA بواسطة الإكسوسوم من قبل مركب ترامب، الذي يحتوي على كل من RNA هيليكاز (Mtr4) ونشاط التذييل بعديد الأدينيلات (Trf4).[25]
وظيفة
وظيفة أنزيمية
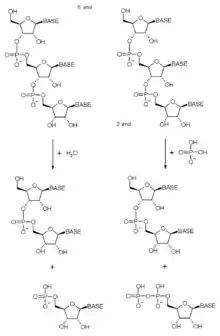
كما ذكر أعلاه، يحتوي مركب إكسوسوم على العديد من البروتينات في نطاقات الريبونوكلياز، وتغيرت الطبيعة الدقيقة لهذه النطاقات من الريبونوكلياز عبر التطور من البكتريا إلى العَتائِق ومنه إلى مركبات حقيقيات النوى حيث تم اكتساب العديد من الأنشطة وفقدها. الإكسوسوم بالدرجة الأولى يكون في إكسوريبونوكلياز 3' - 5'، وهذا يعني أنه يَحُطُّ من جزيئات الحمض النووي الريبوزي (RNA) للنهاية 3'. إكسوريبونوكلياز الوارد في مركبات الإكسوسوم هو إما فوسفوروليتي (مثل بروتينات RNase PH) وفي حقيقيات النوى يكون هدروليتي (مثل بروتينات RNase R و RNase D). تَسْتَخْدِمُ الأنزيمات الفوسفوروليتية الفوسفات غير العضوي لفسخ روابط ثنائي أستر الفوسفور وإطلاق ثنائي فوسفات النوكليوتيد، وتَسْتَخْدِمُ الأنزيمات الهدروليتية الماء لحلمأة هذه الروابط وإطلاق أحادي فوسفات النيوكليوتيد.
وفي العَتائِق، وحدة Rrp41 الفرعية من مركب إكسوريبونوكلياز الفوسفوروليتي، توجد ثلاث نسخ من هذا البروتين في بنية حلقية، وهي مسؤولة عن نشاط المركب.[10] وفي حقيقيات النوى، لم تُحافظ أي من وحدات RNase PH على هذا النشاط التحفيزي، وهذا يعني أن البنية الحلقية الأساسية للإكسوسوم البشري لا تحتوي على بروتين إنزيمي نشط.[26] بالرغم من هذه الخسارة في النشاط التحفيزي، فإن بنية الإكسوسوم الأساسي يتم حفظها بشكل كبير من العَتائِق إلى البشر، مما يشير إلى أن المركب يقوم بوظيفة خلوية حيوية، فَفي حقيقيات النوى، يتم تعويض غياب النشاط الفوسفوروليتي عن طريق وجود الإنزيمات الهيدروليوليكية، المسؤولة عن نشاط ريبونوكلياز الإكسوسوم في مثل هذه الكائنات.[27][28][29]
وكما ذكر أعلاه، ترتبط البروتينات الهدروليتية Rrp6 وRrp44 بالإكسوسوم في الخميرة وفي البشر، بالإضافة إلى Rrp6، هناك بروتينين آخرين، وهما Dis3 وDis3L1 يمكن أن يرتبطان في وضع بروتين الخميرة (Rrp44).[20][21] على الرغم من أن بروتينات المجال S1 كان يعتقد في الأصل أن لها نشاط إكسوريبونوكلياز الفوسفوروليتي 3'-5' أيضًا، فقد تم التساؤل عن وجود هذا النشاط في الآونة الأخيرة وقد يكون لهذه البروتينات دور في الركائز المرتبطة فقط قبل إتلافها بواسطة المركب.[27]
ركائز
ويشارك الإكسوسوم في إتلاف وتجهيز مجموعة واسعة من أنواع الحمض النووي الريبوزي، فَفي سيتوبلازم الخلايا، يُشارك في دوران جزيئات الرنا رسول (mRNA)، ويمكن للمركب أن يحلل جزيئات mRNA التي تم وضع علامة عليها وإتلافها لأنها تحتوي على أخطاء،[30] من خلال تفاعلات مع بروتينات من انحلال متوسط-بدون معنى أو انحلال لا توقفي، وبطريقة بديلة، يتم إتلاف mRNAs كجزء من دورانها الطبيعي، حيث تتفاعل العديد من البروتينات التي تُثَبِّتُ أو تُزَعْزِعُ جزيئات الرنا المرسال من خلال الارتباط بعناصر AU-الغنية في المنطقة 3' غير المترجمة من mRNAs مع مركب الإكسوسوم.[31][32][33] وفي النواة، الإكسوسوم مطلوب لتجهيز العديد من جزيئات RNA الصغيرة النووية بشكل صحيح.[34] وأخيرًا، النوية هي المقصورة التي توجد بها أغلبية مركبات الإكسوسوم الباذخة، ولها دور في معالجة RNA الريبوزومي 5.8S (أول وظيفة محددة من الإكسوسوم) والعديد من الرناوات النووية الصغيرة.[2][34][35]
على الرغم من أن معظم الخلايا تحتوي على إنزيمات أخرى يمكن أن تؤدي إلى إتلاف الحمض النووي الريبوزي (RNA)، إما من النهاية 3' أو من طرف النهاية 5' للحمض النووي الريبوزي (RNA)، فمركب الإكسوسوم ضروري لبقاء الخلية، وعندما يتم تقليل تعبير بروتينات الإكسوسوم بشكل مصطنع أو متوقف، على سبيل المثال عن طريق تداخل RNA، يتوقف النمو وتموت الخلايا في النهاية، كل من البروتينات الأساسية لمركب الإكسوسوم، بالإضافة إلى البروتينين الرئيسيين المقترنين، هي بروتينات أساسية.[36] البكتيريا لا تملك مركب إكسوسوم، ومع ذلك، يتم تنفيذ وظائف مماثلة من قبل مركب أبسط يتضمن بروتين PNPase، ويدعى ديغرادوسوم (degradosome).[37]
الإكسوسوم هو مركب رئيسي في مراقبة جودة الحمض النووي الريبوزي، على عكس حقيقيات النوى، لأن حقيقيات النوى تمتلك أنظمة مراقبة للـ RNA نشطة جدًا والتي تتعرف على مركبات بروتين RNA غير معالجة ومخطئة (مثل الريبوسومات) قبل خروجها من النواة. ومن المفترض أن هذا النظام يمنع المركبات الشاذة من التدخل في العمليات الخلوية الهامة مثل تصنيع البروتين.[38]
بالإضافة إلى معالجة RNA، والدوران وأنشطة الرصد، الإكسوسوم يَكْتَسِبُ أهمية خاصة في إتلاف ما يسمى بالمُنْتسَخَات غير المستقرة الخفية (cryptic unstable transcripts) التي يتم إنتاجها من آلاف المواقع ضمن جينوم الخميرة.[39][40] ولا تزال أهمية هذه الرنا غير المستقرة وإتلافها غير واضحين، ولكن اكتشِفَت أيضًا أنواع RNA مماثلة في الخلايا البشرية.[41]
اضطراب
مناعة ذاتية
مركب الإكسوسوم هو الهدف من الأجسام الذاتية المضادة في المرضى الذين يعانون من أمراض المناعة الذاتية المختلفة، حيث عُثِر على هذه الأجسام المضادة بشكل رئيسي في الأشخاص الذين يعانون من التهاب العضل التصلبي، وهو أحد أمراض المناعة الذاتية التي يعاني منها المرضى بِكُلٍ من تصلب الجلد والتهاب العضلات أو التهاب العضلات والجلد.[42] يمكن الكشف عن الأجسام المضادة في مصل دم المرضى من خلال مجموعة متنوعة من المقايسات. في الماضي، كانت الطرق الأكثر شيوعًا هي الانتشار المناعي المزدوج من خلال استخدام مستخلصات الغدة الزعترية للعجول، أو التَّأَلُّقٌ المَناعِيّ على خلايا HEp-2 أو الترسيب المناعي لمستخلصات الخلايا البشرية. وفي مقايسات الترسيب المناعي مع مصل من الأمصال الإيجابية المضادة للإكسوسوم، حيث تَرَسَّبَت مجموعة مميزة من البروتينات، بالفعل قبل سنوات من تحديد مركب الإكسوسوم، كان يطلق على هذا النمط مصطلح (مركب PM/Scl).[43] يُظْهِرُ التألق المناعي، باستخدام أمصال من هؤلاء المرضى، عادة تلطيخ نموذجي لنويات الخلايا، مما أثار الاقتراح بأن المستضد المعترف به بواسطة الأجسام المضادة الذاتية قد يكون مهمًا في اصطناع الريبوسوم.[44] وفي الآونة الأخيرة، أصبحت بروتينات الإكسوسوم التي يعاد تركيبها متاحة، وقد اسْتُخْدِمَت هذه البروتينات في التنمية المناعية الخطية (LIAs) والمقايسات الامتصاصية المناعية المرتبطة بالإنزيم (ELISAs) لاكتشاف هذه الأجسام المضادة.[8]
وفي هذه الأمراض، يتم توجيه الأجسام المضادة بشكل رئيسي ضد بروتينين من بروتينات المركب، يسميان PM/Scl-100 (مثل بروتين RNase D) و PM/Scl-75 (RNase PH واحد مثل بروتينات الحلقة) وعُثِر على الأجسام المضادة التي تُدْرِكُ هذه البروتينات في ما يقارب 30٪ من المرضى الذين يعانون من التهاب العضل التصلبي.[45] رغم أن هذان البروتينان هما الهدف الرئيسي للأجسام المضادة الذاتية، ويمكن استهداف وحدات فرعية أخرى للإكسوسوم والبروتينات المرتبطة به (مثل C1D) في هؤلاء المرضى.[46][47] وفي الوقت الحالي، الطريقة الأكثر حساسية للكشف عن هذه الأجسام المضادة هي استخدام الببتيد، المشتق من بروتين PM/Scl-100، كمستضد في ELISA، بدلاً من البروتينات الكاملة، وبهذه الطريقة، يُعثَر على الأجسام المضادة في ما يصل إلى 55٪ من المرضى الذين يعانون من التهاب العضل التصلبي، ولكن يمكن أيضا الكشف عنها في المرضى الذين يعانون إما من تصلب الجلد، أو التهاب العضلات، أو التهاب الجلد والعظام فقط.[48]
بما أن الأجسام المضادة توجد بشكل رئيسي في المرضى الذين يعانون من أعراض العديد من أمراض المناعة الذاتية المختلفة، يمكن أن تختلف الأعراض السريرية لهؤلاء المرضى بشكل كبير. الأعراض التي تَظْهَر في أغلب الأحيان هي الأعراض النمطية لأمراض المناعة الذاتية الفردية وتشمل ظاهرة رينود، والتهاب المفاصل، والتهاب العضل وتصلب الجلد.[49] علاج هؤلاء المرضى يكون مَصْحُوب بأَعْرَاض وهو مشابه لعلاج مرض المناعة الذاتية الفردية، وغالبا ما ينطوي إما على الأدوية الكابتة للمناعة أو المُعَدِّلة للمناعة.[50]
علاج السرطان
وقد ثبت أن الإكسوسوم يُثَبَّطُ بواسطة مضاد المئيضة والفلورويوراسيل وهو دواء يستخدم في العلاج الكيميائي للسرطان، وهو واحد من أنجح الأدوية لعلاج الأورام الصلبة. وفي خلايا الخميرة المعالجة بالفلورويوراسيل، عُثِر على عيوب في معالجة الحمض النووي الريبوزي الريبوزومي مماثلة لتلك التي شوهدت عندما تم حظر نشاط الإكسوسوم من خلال إستراتيجيات علم الأحياء الجزيئي. عدم وجود معالجة صحيحة للحمض النووي الريبوزي الريبوزومي يؤدي إلى قتل الخلايا، موضحاً تأثير مضاد المئيضة على الدواء.[51]
اضطرابات عصبية
تُسَبِّبُ الطفرات في عنصر إكسوسوم 3، مرض العصبون الحركي الشوكي الطفيلي، وضمور المخيخ، وصغر الرأس التقدمي وتأخر النمو العميق العالمي، بما يتفق مع نقص تنسج الشرايين من نوع 1B (PCH1B; MIM 614678).[52]
قائمة الوحدات الفرعية
| الرقم | الاسم العام | المجالات | الإنسان | الخميرة (S. cerevisiae) | العتائق | MW (kD) | الجينات البشرية | جينات الخميرة |
|---|---|---|---|---|---|---|---|---|
| 1 | Csl4 | S1 RBD | hCsl4 | Csl4p/Ski4p | Csl4 | 21-32 | EXOSC1 | YNL232W |
| 2 | Rrp4 | S1/KH RBD | hRrp4 | Rrp4p | Rrp4 | 28-39 | EXOSC2 | YHR069C |
| 3 | Rrp40 | S1/KH RBD | hRrp40 | Rrp40p | (Rrp4)A | 27-32 | EXOSC3 | YOL142W |
| 4 | Rrp41 | RNase PH | hRrp41 | Rrp41p/Ski6p | Rrp41C | 26-28 | EXOSC4 | YGR195W |
| 5 | Rrp46 | RNase PH | hRrp46 | Rrp46p | (Rrp41)A,C | 25-28 | EXOSC5 | YGR095C |
| 6 | Mtr3 | RNase PH | hMtr3 | Mtr3p | (Rrp41)A,C | 24-37 | EXOSC6 | YGR158C |
| 7 | Rrp42 | RNase PH | hRrp42 | Rrp42p | Rrp42 | 29-32 | EXOSC7 | YDL111C |
| 8 | Rrp43 | RNase PH | OIP2 | Rrp43p | (Rrp42)A | 30-44 | EXOSC8 | YCR035C |
| 9 | Rrp45 | RNase PH | PM/Scl-75 | Rrp45p | (Rrp42)A | 34-49 | EXOSC9 | YDR280W |
| 10 | Rrp6 | RNase D | PM/Scl-100C | Rrp6pC | n/a | 84-100 | EXOSC10 | YOR001W |
| 11 | Rrp44 | RNase R | Dis3B,C | Rrp44p/Dis3pC | n/a | 105-113 | DIS3
DIS3L1 |
YOL021C |
- A يوجد في العَتائِق مجموعة من بروتينات الإكسوسوم موجودة في نسخ متعددة، لتشكيل الأساس الكامل لمركب الإكسوسوم.
- B في البشر، يمكن ربط بروتينان مختلفان في هذا الموضع. في سيتوبلازم الخلايا، يرتبط Dis3L1 بالإكسوسوم، بينما في النواة، يمكن للـ Dis3 أن يرتبط بالمركب الأساسي.
- C يساهم في نشاط ريبونليوليتيك المركب.
انظر أيضا
- البروتيزوم، الآلية الرئيسية في الخلية لتحلل البروتين.
- جسيم التضفير، وهو مركب يشارك في ربط RNA، ويحتوي أيضا على بنية حلقية لربط الحمض النووي الريبوزي
هوامش
مـ1 (بالإنجليزية: exosome complex (أو PM/Scl complex، وغالبا ما يطلق عليه exosome))
المراجع
- Team, Almaany. "ترجمة و معنى complex بالعربي في قاموس المعاني. قاموس عربي انجليزي مصطلحات صفحة 1". www.almaany.com (باللغة الإنجليزية). مؤرشف من الأصل في 25 سبتمبر 2018. اطلع عليه بتاريخ 24 سبتمبر 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Mitchell, P; Petfalski, E; Shevchenko, A; Mann, M; Tollervey, D (1997). "The Exosome: A Conserved Eukaryotic RNA Processing Complex Containing Multiple 3′→5′ Exoribonucleases". Cell. 91 (4): 457–466. doi:10.1016/S0092-8674(00)80432-8. PMID 9390555. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Allmang, C; Petfalski, E; Podtelejnikov, A; Mann, M; Tollervey, D; Mitchell, P (1999). "The yeast exosome and human PM-Scl are related complexes of 3' --> 5' exonucleases". Genes & Development. 13 (16): 2148–58. doi:10.1101/gad.13.16.2148. PMC 316947. PMID 10465791. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Brouwer, R; Allmang, C; Raijmakers, R; Van Aarssen, Y; Egberts, WV; Petfalski, E; Van Venrooij, WJ; Tollervey, D; Pruijn, GJ (2001). "Three novel components of the human exosome". Journal of Biological Chemistry. 276 (9): 6177–84. doi:10.1074/jbc.M007603200. PMID 11110791. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Chen, CY; Gherzi, R; Ong, SE; Chan, EL; Raijmakers, R; Pruijn, GJ; Stoecklin, G; Moroni, C; et al. (2001). "AU binding proteins recruit the exosome to degrade ARE-containing mRNAs". Cell. 107 (4): 451–64. doi:10.1016/S0092-8674(01)00578-5. PMID 11719186. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Koonin, EV; Wolf, YI; Aravind, L (2001). "Prediction of the archaeal exosome and its connections with the proteasome and the translation and transcription machineries by a comparative-genomic approach". Genome Research. 11 (2): 240–52. doi:10.1101/gr.162001. PMC 311015. PMID 11157787. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Evguenieva-Hackenberg, E; Walter, P; Hochleitner, E; Lottspeich, F; Klug, G (2003). "An exosome-like complex in Sulfolobus solfataricus". EMBO Reports. 4 (9): 889–93. doi:10.1038/sj.embor.embor929. PMC 1326366. PMID 12947419. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Schilders, G; Van Dijk, E; Raijmakers, R; Pruijn, GJ (2006). "Cell and molecular biology of the exosome: how to make or break an RNA". International review of cytology. 251: 159–208. doi:10.1016/S0074-7696(06)51005-8. ISBN 9780123646552. PMID 16939780. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lorentzen, E; Walter, P; Fribourg, S; Evguenieva-Hackenberg, E; Klug, G; Conti, E (2005). "The archaeal exosome core is a hexameric ring structure with three catalytic subunits". Nature Structural & Molecular Biology. 12 (7): 575–81. doi:10.1038/nsmb952. PMID 15951817. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Shen, V; Kiledjian, M (2006). "A view to a kill: structure of the RNA exosome". Cell. 127 (6): 1093–5. doi:10.1016/j.cell.2006.11.035. PMC 1986773. PMID 17174886. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Raijmakers, R; Egberts, WV; Van Venrooij, WJ; Pruijn, GJ (2002). "Protein-protein interactions between human exosome components support the assembly of RNase PH-type subunits into a six-membered PNPase-like ring". Journal of Molecular Biology. 323 (4): 653–63. doi:10.1016/S0022-2836(02)00947-6. PMID 12419256. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Walter, P; Klein, F; Lorentzen, E; Ilchmann, A; Klug, G; Evguenieva-Hackenberg, E (2006). "Characterization of native and reconstituted exosome complexes from the hyperthermophilic archaeon Sulfolobus solfataricus". Molecular Microbiology. 62 (4): 1076–89. doi:10.1111/j.1365-2958.2006.05393.x. PMID 17078816. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ishii, R; Nureki, O; Yokoyama, S (2003). "Crystal structure of the tRNA processing enzyme RNase PH from Aquifex aeolicus". Journal of Biological Chemistry. 278 (34): 32397–404. doi:10.1074/jbc.M300639200. PMID 12746447. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Symmons, MF; Jones, GH; Luisi, BF (2000). "A duplicated fold is the structural basis for polynucleotide phosphorylase catalytic activity, processivity, and regulation". Structure. 8 (11): 1215–26. doi:10.1016/S0969-2126(00)00521-9. PMID 11080643. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lin-Chao, S; Chiou, NT; Schuster, G (2007). "The PNPase, exosome and RNA helicases as the building components of evolutionarily-conserved RNA degradation machines". Journal of Biomedical Science. 14 (4): 523–32. doi:10.1007/s11373-007-9178-y. PMID 17514363. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Harlow, LS; Kadziola, A; Jensen, KF; Larsen, S (2004). "Crystal structure of the phosphorolytic exoribonuclease RNase PH from Bacillus subtilis and implications for its quaternary structure and tRNA binding". Protein Science. 13 (3): 668–77. doi:10.1110/ps.03477004. PMC 2286726. PMID 14767080. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lebreton, A; Tomecki, R; Dziembowski, A; Séraphin, B (2008). "Endonucleolytic RNA cleavage by a eukaryotic exosome". Nature. 456 (7224): 993–6. Bibcode:2008Natur.456..993L. doi:10.1038/nature07480. PMID 19060886. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Schneider, C; Leung, E; Brown, J; Tollervey, D (2009). "The N-terminal PIN domain of the exosome subunit Rrp44 harbors endonuclease activity and tethers Rrp44 to the yeast core exosome". Nucleic Acids Research. 37 (4): 1127–40. doi:10.1093/nar/gkn1020. PMC 2651783. PMID 19129231. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Schneider, C; Anderson, JT; Tollervey, D (2007). "The exosome subunit Rrp44 plays a direct role in RNA substrate recognition". Molecular Cell. 27 (2): 324–31. doi:10.1016/j.molcel.2007.06.006. PMID 17643380. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Staals, RH; Bronkhorst, AW; Schilders, G; Slomovic, S; Schuster, G; Heck, AJ; Raijmakers, R; Pruijn, GJ (2010). "Dis3-like 1: a novel exoribonuclease associated with the human exosome". The EMBO Journal. 29 (14): 2358–67. doi:10.1038/emboj.2010.122. PMC 2910272. PMID 20531389. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Tomecki, R; Kristiansen, MS; Lykke-Andersen, S; Chlebowski, A; Larsen, KM; Szczesny, RJ; Drazkowska, K; Pastula, A; et al. (2010). "The human core exosome interacts with differentially localized processive RNases: hDIS3 and hDIS3L". The EMBO Journal. 29 (14): 2342–57. doi:10.1038/emboj.2010.121. PMC 2910271. PMID 20531386. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Mian, IS (1997). "Comparative sequence analysis of ribonucleases HII, III, II PH and D". Nucleic Acids Research. 25 (16): 3187–3195. doi:10.1093/nar/25.16.3187. PMC 146874. PMID 9241229. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Raijmakers, R; Schilders, G; Pruijn, GJ (2004). "The exosome, a molecular machine for controlled RNA degradation in both nucleus and cytoplasm". European Journal of Cell Biology. 83 (5): 175–83. doi:10.1078/0171-9335-00385. PMID 15346807. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wang, L; Lewis, MS; Johnson, AW (2005). "Domain interactions within the Ski2/3/8 complex and between the Ski complex and Ski7p". RNA. 11 (8): 1291–302. doi:10.1261/rna.2060405. PMC 1370812. PMID 16043509. الوسيط
|CitationClass=تم تجاهله (مساعدة) - LaCava, J; Houseley, J; Saveanu, C; Petfalski, E; Thompson, E; Jacquier, A; Tollervey, D (2005). "RNA degradation by the exosome is promoted by a nuclear polyadenylation complex". Cell. 121 (5): 713–24. doi:10.1016/j.cell.2005.04.029. PMID 15935758. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Liu, Q; Greimann, JC; Lima, CD (2007). "Erratum: Reconstitution, activities, and structure of the eukaryotic RNA exosome". Cell. 131 (1): 188–189. doi:10.1016/j.cell.2007.09.019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Dziembowski, A; Lorentzen, E; Conti, E; Séraphin, B (2007). "A single subunit, Dis3, is in essence responsible for yeast exosome core activity". Nature Structural & Molecular Biology. 14 (1): 15–22. doi:10.1038/nsmb1184. PMID 17173052. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Liu, Q; Greimann, JC; Lima, CD (2006). "Reconstitution, activities, and structure of the eukaryotic RNA exosome". Cell. 127 (6): 1223–37. doi:10.1016/j.cell.2006.10.037. PMID 17174896. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lorentzen, E; Conti, E (2005). "Structural basis of 3' end RNA recognition and exoribonucleolytic cleavage by an exosome RNase PH core". Molecular Cell. 20 (3): 473–81. doi:10.1016/j.molcel.2005.10.020. PMID 16285928. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "«الإكسوسوم» يحضِّر لقضم الحمض النووي الريبِي". arabicedition.nature.com. مؤرشف من الأصل في 27 نوفمبر 2015. اطلع عليه بتاريخ 19 سبتمبر 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - LeJeune, F; Li, X; Maquat, LE (2003). "Nonsense-mediated mRNA decay in mammalian cells involves decapping, deadenylating, and exonucleolytic activities". Molecular Cell. 12 (3): 675–87. doi:10.1016/S1097-2765(03)00349-6. PMID 14527413. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wilson, MA; Meaux, S; Van Hoof, A (2007). "A genomic screen in yeast reveals novel aspects of nonstop mRNA metabolism". Genetics. 177 (2): 773–84. doi:10.1534/genetics.107.073205. PMC 2034642. PMID 17660569. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lin, WJ; Duffy, A; Chen, CY (2007). "Localization of AU-rich element-containing mRNA in cytoplasmic granules containing exosome subunits". Journal of Biological Chemistry. 282 (27): 19958–68. doi:10.1074/jbc.M702281200. PMID 17470429. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Allmang, C; Kufel, J; Chanfreau, G; Mitchell, P; Petfalski, E; Tollervey, D (1999). "Functions of the exosome in rRNA, snoRNA and snRNA synthesis". EMBO Journal. 18 (19): 5399–410. doi:10.1093/emboj/18.19.5399. PMC 1171609. PMID 10508172. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Schilders, G; Raijmakers, R; Raats, JM; Pruijn, GJ (2005). "MPP6 is an exosome-associated RNA-binding protein involved in 5.8S rRNA maturation". Nucleic Acids Research. 33 (21): 6795–804. doi:10.1093/nar/gki982. PMC 1310903. PMID 16396833. الوسيط
|CitationClass=تم تجاهله (مساعدة) - van Dijk, EL; Schilders, G; Pruijn, GJ (2007). "Human cell growth requires a functional cytoplasmic exosome, which is involved in various mRNA decay pathways". RNA. 13 (7): 1027–35. doi:10.1261/rna.575107. PMC 1894934. PMID 17545563. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Carpousis AJ, AJ (2002). "The Escherichia coli RNA degradosome: structure, function and relationship in other ribonucleolytic multienzyme complexes". Biochem. Soc. Trans. 30 (2): 150–5. doi:10.1042/BST0300150. PMID 12035760. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Houseley J, LaCava J, Tollervey D (يوليو 2006). "RNA-quality control by the exosome". Nat. Rev. Mol. Cell Biol. 7 (7): 529–39. doi:10.1038/nrm1964. PMID 16829983. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wyers F, F; Rougemaille, M; Badis, G; et al. (يونيو 2005). "Cryptic pol II transcripts are degraded by a nuclear quality control pathway involving a new poly(A) polymerase". Cell. 121 (5): 725–37. doi:10.1016/j.cell.2005.04.030. PMID 15935759. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Neil H, Malabat C, d'Aubenton-Carafa Y, Xu Z, Steinmetz LM, Jacquier A (فبراير 2009). "Widespread bidirectional promoters are the major source of cryptic transcripts in yeast". Nature. 457 (7232): 1038–42. Bibcode:2009Natur.457.1038N. doi:10.1038/nature07747. PMID 19169244. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Preker P, P; Nielsen, J; Kammler, S; et al. (ديسمبر 2008). "RNA exosome depletion reveals transcription upstream of active human promoters". Science. 322 (5909): 1851–4. Bibcode:2008Sci...322.1851P. doi:10.1126/science.1164096. PMID 19056938. الوسيط
|CitationClass=تم تجاهله (مساعدة) - J.E. Pope, JE (2002). "Scleroderma overlap syndromes". Current Opinion in Rheumatology. 14 (6): 704–10. doi:10.1097/00002281-200211000-00013. PMID 12410095. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Gelpi, C; Algueró, A; Angeles Martinez, M; Vidal, S; Juarez, C; Rodriguez-Sanchez, JL (1991). "Identification of protein components reactive with anti-PM/Scl autoantibodies". Clinical and Experimental Immunology. 81 (1): 59–64. doi:10.1111/j.1365-2249.1990.tb05291.x. PMC 1535032. PMID 2199097. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Targoff, IN; Reichlin, M (1985). "Nucleolar localization of the PM-Scl antigen". Arthritis & Rheumatism. 28 (2): 226–30. doi:10.1002/art.1780280221. PMID 3918546. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Raijmakers, R; Renz, M; Wiemann, C; Egberts, WV; Seelig, HP; Van Venrooij, WJ; Pruijn, GJ (2004). "PM-Scl-75 is the main autoantigen in patients with the polymyositis/scleroderma overlap syndrome". Arthritis & Rheumatism. 50 (2): 565–9. doi:10.1002/art.20056. PMID 14872500. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Brouwer, R; Vree Egberts, WT; Hengstman, GJ; Raijmakers, R; Van Engelen, BG; Seelig, HP; Renz, M; Mierau, R; et al. (2002). "Autoantibodies directed to novel components of the PM/Scl complex, the human exosome". Arthritis Research & Therapy. 4 (2): 134–8. doi:10.1186/ar389. PMC 83843. PMID 11879549. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Schilders, G; Egberts, WV; Raijmakers, R; Pruijn, GJ (2007). "C1D is a major autoantibody target in patients with the polymyositis-scleroderma overlap syndrome". Arthritis & Rheumatism. 56 (7): 2449–54. doi:10.1002/art.22710. PMID 17599775. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Mahler, M; Raijmakers, R; Dähnrich, C; Blüthner, M; Fritzler, MJ (2005). "Clinical evaluation of autoantibodies to a novel PM/Scl peptide antigen". Arthritis Research & Therapy. 7 (3): R704–13. doi:10.1186/ar1729. PMC 1174964. PMID 15899056. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Mahler, M; Raijmakers, R (2007). "Novel aspects of autoantibodies to the PM/Scl complex: Clinical, genetic and diagnostic insights". Autoimmunity Reviews. 6 (7): 432–7. doi:10.1016/j.autrev.2007.01.013. PMID 17643929. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Jablonska, S; Blaszczyk, M (1998). "Scleromyositis: a scleroderma/polymyositis overlap syndrome". Clinical Rheumatology. 17 (6): 465–7. doi:10.1007/BF01451281. PMID 9890673. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lum, PY; Armour, CD; Stepaniants, SB; Cavet, G; Wolf, MK; Butler, JS; Hinshaw, JC; Garnier, P; et al. (2004). "Discovering modes of action for therapeutic compounds using a genome-wide screen of yeast heterozygotes". Cell. 116 (1): 121–37. doi:10.1016/S0092-8674(03)01035-3. PMID 14718172. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wan, J.; Yourshaw, M.; Mamsa, H.; Rudnik-Schöneborn, S.; Menezes, M. P.; Hong, J. E.; Leong, D. W.; Senderek, J.; Salman, M. S.; Chitayat, D.; Seeman, P.; Von Moers, A.; Graul-Neumann, L.; Kornberg, A. J.; Castro-Gago, M.; Sobrido, M. A. J. S.; Sanefuji, M.; Shieh, P. B.; Salamon, N.; Kim, R. C.; Vinters, H. V.; Chen, Z.; Zerres, K.; Ryan, M. M.; Nelson, S. F.; Jen, J. C. (2012). "Mutations in the RNA exosome component gene EXOSC3 cause pontocerebellar hypoplasia and spinal motor neuron degeneration". Nature Genetics. 44 (6): 704–708. doi:10.1038/ng.2254. PMC 3366034. PMID 22544365. الوسيط
|CitationClass=تم تجاهله (مساعدة)
كتب
- Schilders, G; Pruijn, GJ (2008). "Biochemical studies of the mammalian exosome with intact cells". Methods Enzymol. 448: 211–226. doi:10.1016/S0076-6879(08)02611-6. ISBN 9780123743787. PMID 19111178. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Houseley, J; Tollervey, D (2008). "The nuclear RNA surveillance machinery: the link between ncRNAs and genome structure in budding yeast?". Biochim Biophys Acta. 1779 (4): 239–246. doi:10.1016/j.bbagrm.2007.12.008. PMID 18211833. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Vanacova, S; Stefl, R (2007). "The exosome and RNA quality control in the nucleus". EMBO Reports. 8 (7): 651–657. doi:10.1038/sj.embor.7401005. PMC 1905902. PMID 17603538. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Houseley, J; Lacava, J; Tollervey, D (2006). "RNA-quality control by the exosome". Nature Reviews Molecular Cell Biology. 7 (7): 529–539. doi:10.1038/nrm1964. PMID 16829983. الوسيط
|CitationClass=تم تجاهله (مساعدة) –-- subscription required - Büttner, K; Wenig, K; Hopfner, KP (2006). "The exosome: a macromolecular cage for controlled RNA degradation". Molecular Microbiology. 61 (6): 1372–1379. doi:10.1111/j.1365-2958.2006.05331.x. PMID 16968219. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lorentzen, E; Conti, E (2006). "The Exosome and the Proteasome: Nano-Compartments for Degradation". Cell. 125 (4): 651–654. doi:10.1016/j.cell.2006.05.002. PMID 16713559. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Pruijn, GJ (2005). "Doughnuts dealing with RNA". Nature Structural & Molecular Biology. 12 (7): 562–564. doi:10.1038/nsmb0705-562. PMID 15999107. الوسيط
|CitationClass=تم تجاهله (مساعدة)
وصلات خارجية
- هيكل إكسوسوم الإنسان في بنك معلومات بروتين RCSB
- هيكل إكسوسوم العَتائِق في بنك معلومات بروتين RCSB
- هيكل إكسوسوم العَتائِق لربط RNA في بنك معلومات بروتين RCSB
- هيكل إكسوسوم بروتين الخميرة Rrp6 في بنك معلومات بروتين RCSB
- هياكل جزيئية ثلاثية الأبعاد للإكسوسوم في بنك معلومات EM (EMDB)
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علم الأحياء
بوابة علم الأحياء بوابة تقانة حيوية
بوابة تقانة حيوية بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة طب
بوابة طب