مركب إسكات مستحث بالرنا
مركب إسكات مُسْتَحَثْ بالرنا (بالإنجليزية: RNA-induced silencing complex) واختصارا (ريسك : RISC) هو مركب بروتيني عديد البروتينات يتكون من بروتين من عائلة الأرغنوت بالإضافة إلى بروتينات نووية ريبوزية أخرى، والتي تقوم بالارتباط بسلسلة رنا مفردة مثل الرنا الميكروي أو الرنا المتدخل الصغير عالجها سابقا النوكلياز الداخلي دايسر.[1]
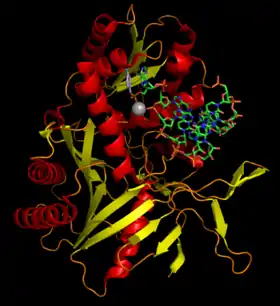
يقص دايسر سلسلة الرنا المتدخل الصغير الأولي المزدوجة ويُنتِج سلسلتين مفردتين، واحدة منهما فقط يتم إدراجها في مركب ريسك والأخرى يتم تفكيكيها. بعدها تعمل تلك السلسلة كقالب لمركب ريسك يتعرف من خلالها على سلاسل الرنا الرسول المكملة وحين يجدها يتم تثبيط النسخ إن كان التكامل بينهما غير تام، أما إن كان تاما أو شبه تام فإن مركب ريسك يفعّل بروتين الأرغنوت الذي يقص الرنا الرسول في مركز الترابط بواسطة نطاق بيوي الخاص به.
تسمى هذه العملية تدخل الرنا وتتواجد لدى حقيقيات النوى، وهي عملية مفتاحية في إسكات الجينات والدفاع ضد عدوى الفيروسات.[2][3]
الارتباط بالرنا الرسول
ليس واضحا لحد الآن كيف يحدد مركب ريسك المنشَّط جزيئات الرنا المستهدفة في الخلية، رغم أنه تبين أن هذه العملية تحدث حتى في الحالات التي لا يكون فيها الرنا الرسول خاضعا للترجمة لتخليق بروتين.[4]
جزيئات الرنا الميكروي المعبر عنها ذاتيا لدى الحيوانات ليست في العادة متكاملة بشكل تام مع الكثير من جزيئات الرنا الرسول، وتقوم بتعديل التعبير الجيني عبر تثبيط الترجمة.[5][6] أما عند النبات فعملية الارتباط أكثر دقة وتكاملا مع جزيئات الرنا الرسول وفي العادة كل رنا ميكروي يرتبط مع رنا رسول واحد فقط. الدقة الأكثر تعني أن تفكيك الرنا الرسول على الأرجح سيحدث.[7]
وظيفته في تدخل الرنا
تحميل الرنا مزدوج السلاسل
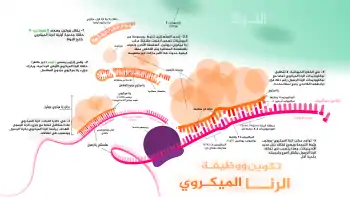
يساعد الرناز 3 دايسر مركب ريسك في تدخل الرنا عبر قص الرنا مزدوج السلاسل إلى قطعة طولها 21-23 نوكليوتيد مع فارق نوكليوتيدتين في النهاية 3'.[8][9] بعد ذلك تُدرج هاتين القطعتين في مركب ريسك ويكون لهما مصير مختلف حسب ظاهرة قانون اللاتناظر.[10][11][12]
- يختار بروتين الأرغنوت السلسلة الأقل استقرارا في النهاية 5' ويُدرجها في مركب ريسك.[12][13] تعرف هذه السلسلة بالسلسلة المُرشِدة.
- السلسلة الأخرى التي تعرف باسم السلسلة المسافرة، يتم تفكيكها بواسطة ريسك.[14]
المراجع
- Filipowicz W, Bhattacharyya SN, Sonenber N (2008). "Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight?". Nature Reviews Genetics. 9 (2): 102–114. doi:10.1038/nrg2290. PMID 18197166. الوسيط
|CitationClass=تم تجاهله (مساعدة) -
Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC (1998). "Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans". نيتشر. 391 (6669): 806–811. doi:10.1038/35888. PMID 9486653. الوسيط
|CitationClass=تم تجاهله (مساعدة) -
Watson, James D. (2008). Molecular Biology of the Gene. San Francisco, CA: Cold Spring Harbor Laboratory Press. صفحات 641–648. ISBN 978-0-8053-9592-1. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Sen GL, Wehrman TS, Blau HM (2005). "mRNA translation is not a prerequisite for small interfering RNA-mediated mRNA cleavage". Differentiation. 73 (6): 287–293. doi:10.1111/j.1432-0436.2005.00029. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Saumet A, Lecellier CH (2006). "Anti-viral RNA silencing: do we look like plants?". Retrovirology. 3: 3. doi:10.1186/1742-4690-3-3. PMC 1363733. PMID 16409629. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bartel DP (2009). "MicroRNAs: target recognition and regulatory functions". Cell. 136 (2): 215–233. doi:10.1016/j.cell.2009.01.002. PMC 3794896. PMID 19167326. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Jones-Rhoades MW, Bartel DP, Bartel B (2006). "MicroRNAs and their regulator roles in plants". Annual Review of Plant Biology. 57: 19–53. doi:10.1146/annurev.arplant.57.032905.105218. PMID 16669754. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Zamore PD, Tuschl T, Sharp PA, Bartel DP (2000). "RNAi: double-stranded RNA directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide intervals". Cell. 101 (1): 25–33. doi:10.1016/S0092-8674(00)80620-0. PMID 10778853. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Vermeulen A, Behlen L, Reynolds A, Wolfson A, Marshall W, Karpilow J, Khvorova A (2005). "The contributions of dsRNA structure to Dicer specificity and efficiency". RNA. 11 (5): 674–682. doi:10.1261/rna.7272305. PMC 1370754. PMID 15811921. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Schwarz DS, Hutvágner G, Du T, Xu Z, Aronin N, Zamore PD (2003). "Asymmetry in the assembly of the RNAi enzyme complex". Cell. 115 (2): 199–208. doi:10.1016/S0092-8674(03)00759-1. PMID 14567917. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Khvorova A, Reynolds A, Jayasena SD (2003). "Functional siRNAs and miRNAs exhibit strand bias". Cell. 115 (2): 209–216. doi:10.1016/S0092-8674(03)00801-8. PMID 14567918. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Siomi H, Siomi MC (2009). "On the road to reading the RNA-interference code". نيتشر. 457 (7228): 396–404. doi:10.1038/nature07754. PMID 19158785. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Preall JB, He Z, Gorra JM, Sontheimer EJ (2006). "Short interfering RNA strand selection is independent of dsRNA processing polarity during RNAi in Drosophila". Current Biology. 16 (5): 530–535. doi:10.1016/j.cub.2006.01.061. PMID 16527750. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Gregory RI, Chendrimada TP, Cooch N, Shiekhattar R (2005). "Human RISC couples microRNA biogenesis and posttranscriptional gene silencing". Cell. 123 (4): 631–640. doi:10.1016/j.cell.2005.10.022. PMID 16271387. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي
