وصل بديل
الوصل البديل[1] أو التضفير البديل (بالإنجليزية: Alternative splicing)، هي عملية مُنظمة تحدث أثناء التعبير الجيني حيث بإمكان نسخة أوليّة واحدة أن تُنتج نُسخاً مختلفة من حمض نووي ريبوزي رسول من خلال عملية الوصل؛ حيث يتم ضم أطقم مُختلفة من التعاقدات المُشفرة في كل منها. وهي تُعتبر ظاهرة طبيعية في حقيقيات النوى.
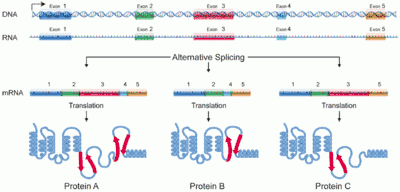
تُشير الأدلة إلى أن النمط الطبيعي لعملية الوصل مُهمة لوظائف المخلوقات. وقد قُدر أن 15% من الاضطرابات الوراثية في الإنسان يرجع سببها إلى تغيير في عملية الوصل. فبإمكان طفرات الوصل أن مواقع جديدة للوصل أو تُلفي النمط الطبيعي له. وهذا يؤدي إلى حدوث خلل وراثي له.
اكتشافها
أوّل ما لوحظت عملية الوصل المتبادل في عام 1977،[2] إذ تُنتج الفيروسات الغدانية خمس نسخ أولية في دورتها الإنتانية قبل انتساخ الحمض النووي الفيروسي، ونسخة أخرى لاحقًا بعد بدء الانتساخ، ويستمر إنتاج هذه النسخ خلال الدورة، ويكون عدد النسخ الإضافية الأولية التي تُنتج في نهاية الدورة كبير، وأكثر بكثير مما يمكن أن يمثله حمض نووي ريبوزي رسول في خلية واحدة. [3][4][5]
وجد الباحثون أن نسخة الحمض النووي الريبوزي الأولية التي تنتجها الفيروسات الغدانية نمط 2 في الطور الأخير تتصل بطرق عديدة مختلفة، مؤدية بذلك إلى إنتاج أحماض نووية ريبوزية رسولة عديدة ترمّز بروتينات فيروسية مختلفة، واحتوت هذه النسخ الأولية على عدة مناطق تحدث فيها عملية تذييل بعديد الأدينيلات، مؤدية للحصول على نهايات 3' مختلفة بدءًا من رنا مرسال واحد.[6]
في عام 1981 وُصف أول مثال للوصل المتبادل في نسخة من جين طبيعي داخلي المنشأ، فقد وجد أن الجين الذي يرمّز الهرمون الدرقي الكالسيتونين يقوم بعملية وصل متبادل في خلايا الثدييات، إذ تحتوي النسخة الأولية من هذا الجين على 6 إكسونات، يحتوي الرنا المرسال المسؤول عن تصنيع الكالسيتونين على الإكسونات من 1 إلى 4، وينتهي بعد تذييل الموقع بعديد الأدينيلات في الإكسون الرابع، ويُنتَج رنا مرسال آخر منه عبر تخطّي الإكسون الرابع، ويكون بذلك يحتوي على الإكسونات من 1 إلى 3 وَ5 وَ6، وهو يرمّز بروتين يُعرف باسم سي جي أر بي (الببتيد المرتبط بجين الكالسيتونين).[7][8]
لوحظ أيضًا بعض الأمثلة على الوصل المتبادل في النُّسخ التي ينتجها الجين المسؤول عن ترميز الغلوبولينات المناعية في الثدييات في أوائل ثمانينيات القرن العشرين.
ومنذ ذلك الحين لوحظ بأن عملية الوصل المتبادل منتشرة في كل مكان في حقيقيات النوى، وحقق جين يدعى Dscam موجود عند ذبابة الفاكهة الشائعة الرقم القياسي حتى الآن في عدد عمليات الوصل المتبادل التي يجريها، فهو قادر على إنتاج 38016 نسخة وصلية مختلفة.[9]
الأنماط
صُنّفت خمسة أنماط رئيسية من أنماط الوصل المتبادل هي:[10]
- ترك أو تخطّي الإكسون: في هذه الحالة قد ينفصل إكسون عن النسخة الأولية، وقد يُحتفظ به، وهو النمط الأشيع عند الثدييات في إنتاج الرنا المرسال الأولي.
- تبادل الأكسونات الحصرية: يُحتفَظ بأحد الإكسونان في الرنا المرسال بعد عملية الوصل، لكن ليس بكِلا الإكسونين.
- تبديل الموقع المانح: يُستخدم موقع وصل بديل هو الوصلة 5' (الموقع المانح) بدلًا من الحدّ 3' في الإكسون المقابل.
- تبديل الموقع القابِل: يُستخدم موقع وصل بديل هو 3' (الموقع القابِل) بدلًا من الحدّ 5' في الإكسون الموافق.
- الإبقاء على الإنترون: يُمكن فصل سلسلة الإنترون، أو ببساطة الإبقاء عليها، ويختلف هذا عن الاحتفاظ بالإكسون لأن السلسلة الباقية لا تكون مُحاطة بأي إنترون، ويكون الإنترون الباقي في منطقة الترميز، ولذلك سيرمز أحماض أمينية كما تفعل الإكسونات المجاورة، أو أنه سيشكل كود التوقف عن النسخ، أو أنه سيسبب تغير في النمط الناتج، ويؤدي بذلك إلى إنتاج بروتين غير وظيفي.
يُعتبر النمط الأخير الأندر عند الثدييات.[11]
يوجد إضافة لهذه الأنماط الرئيسية لعملية الوصل المتبادل آليتان رئيسيتان أيضًا يمكن من خلالهما إنشاء نسخ مختلفة من الرنا المرسال من نفس الجين هما المعزّزات المتعددة ومواقع التذييل بعديد الأدينيلات المختلفة.
تصف هذه النماذج آليات الوصل المتبادل الأساسية، لكنها غير كافية لوصف العمليات المعقدة التي تحدث خلال عملية الوصل المتبادل.
الآلية
الآليات العامة لعملية الوصل المتبادل
عندما يُنتسخ الحمض النووي الريبي الأولي (الرنا المرسال الأولي) من الحمض النووي (الدي إن آي) سيحتوي على العديد من الإنترونات، والأكسونات (يكون المتوسط عند الديدان الخيطية 4 إلى 5 إكسونات وإنترونات، وفي ذبابة الفاكهة أكثر من 100 إنترون وإكسون في رنا مرسال أولي واحد فقط)، وتُحدّد الإكسونات التي سيُحتفَظ بها من خلال عملية الوصل المتبادل، والمسؤول عن تحديد وتنظيم مواقع الوصل هي متممات محفّزات الوصل، وبروتينات الوصل الكاظمة، إضافة للعناصر المقترنة بالرنا المرسال الأولي نفسها.
مَثيلة الحمض النووي الريبي وعملية الوصل المتبادل في الحشرات الاجتماعية
أظهرة مثيلة موقع ثنائي النكليوتيد سي بي جي للحمض النووي دورًا في تنظيم عملية الوصل المتبادل عند الحشرات الاجتماعية، إذ تنظّم هذه المَثيلة عند نحل العسل عملية تخطّي الإكسون بحسب بعض الدراسات الجينية التي أجريت بعد دراسة جينوم هذه الحيوانات.[12]
الأمراض المتعلقة بعملية الوصل المتبادل
أشارت دراسة أجريت في عام 2005 اعتمدت على تحليلات احتمالية بأن أكثر من 60% من الطفرات المسببة للأمراض البشرية تؤثر على عملية الوصل المتبادل، بدلًا من التأثير على سلاسل الترميز، وتشير الدراسات الأحدث إلى أنه قد تتعلّق ثلث الأمراض الوراثية بطريقة ما في عملية الوصل المتبادل، لكن بغض النظر عن النسب المئوية الدقيقة، فإن الأمراض الوراثية المتعلقة بعملية الوصل المتبادل موجودة، والسرطان هو أحد الأمثلة البارزة على الموضوع.[13]
عُثر في نسبة عالية من الخلايا السرطانية على رنا مرسال يقوم بعمليات وصل متبادل غير طبيعية، وكشفت تحليلات تسلسلات الرنا وجود شذوذات في عملية الوصل المتبادل في مسار سرطانات عديدة، لكن من غير المعروف فيما إذا كانت هذه الأنماط الشاذة تسهم في تشكل السرطان، أو أنها مجرد نتيجة للشذوذات الخلوية التي تحدث في سياقه، ولوحظ في أنواع معينة من السرطانات، كسرطانات الكولون، والمستقيم، والبروستات أن الأخطاء في عملية الوصل المتبادل تختلف بشكل كبير بين السرطانات، ويشار إلى هذه الظاهرة بعدم الاستقرار النسخي.[14][15]
يوجد في الواقع نقص في عملية الوصل المتبادل في الخلايا السرطانية مقارنة بالطبيعية، إضافة للاختلافات في أنماط هذه العملية، فمثلًا تحافظ الخلايا السرطانية على مستويات أعلى من الإنترونات، وأقل من الإكسونات، وقد يكون السبب في الاختلافات في عملية الوصل في الخلايا السرطانية هو الطفرات الجسدية في الجينات المسؤولة عن عوامل الوصل، أو التغيرات في فسفرة هذه العوامل داخل الخلايا السرطانية.[14]
تحليل الجينوم واسع الطيف
إن تحليل الجينوم واسع الطيف لعملية الوصل المتبادل مهمة صعبة، لكن تطوّرت الأبحاث التي ستساعد على ذلك، بما فيها التحليلات القائمة على الحمض النووي الميكروبي، والمقايسات المرتبطة بالحمض النووي الريبي، وتقنيات التسلسل العميق، ويمكن استخدام هذه الطرق للبحث عن تعدد الأشكال الظاهرية، والطفرات في أو المتعلقة بعناصر الوصل المتبادل.[16]
استخدمت تقنيات التسلسل العميق لإجراء تحليلات واسعة الطيف على الرنا المرسال (المُعالَج، وغير المُعالَج) مما أعطانا معلومات أكثر عن عملية الوصل المتبادل، فمثلًا تشير نتائج التسلسل العميق إلى أنه عند البشر يخضع تقريبًا 95% من النسخ المكونة من إكسونات متعددة لعملية الوصل المتبادل.[17]
المراجع
- علم الأحياء - بواسطة بيتر هـ. ريفن،جورج ب. جونسون،جوناثان ب. لوسوس،كينيث - صـ 290
- Chow LT, Gelinas RE, Broker TR, Roberts RJ (September 1977). "An amazing sequence arrangement at the 5' ends of adenovirus 2 messenger RNA". Cell. 12 (1): 1–8. doi:10.1016/0092-8674(77)90180-5. PMID 902310. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Leff SE, Rosenfeld MG, Evans RM (1986). "Complex transcriptional units: diversity in gene expression by alternative RNA processing". Annual Review of Biochemistry. 55 (1): 1091–117. doi:10.1146/annurev.bi.55.070186.005303. PMID 3017190. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Chow LT, Broker TR (October 1978). "The spliced structures of adenovirus 2 fiber message and the other late mRNAs". Cell. 15 (2): 497–510. doi:10.1016/0092-8674(78)90019-3. PMID 719751. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Nevins JR, Darnell JE (December 1978). "Steps in the processing of Ad2 mRNA: poly(A)+ nuclear sequences are conserved and poly(A) addition precedes splicing". Cell. 15 (4): 1477–93. doi:10.1016/0092-8674(78)90071-5. PMID 729004. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Maki R, Roeder W, Traunecker A, Sidman C, Wabl M, Raschke W, Tonegawa S (May 1981). "The role of DNA rearrangement and alternative RNA processing in the expression of immunoglobulin delta genes". Cell. 24 (2): 353–65. doi:10.1016/0092-8674(81)90325-1. PMID 6786756. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Rosenfeld MG, Amara SG, Roos BA, Ong ES, Evans RM (March 1981). "Altered expression of the calcitonin gene associated with RNA polymorphism". Nature. 290 (5801): 63–5. Bibcode:1981Natur.290...63R. doi:10.1038/290063a0. PMID 7207587. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Rosenfeld MG, Lin CR, Amara SG, Stolarsky L, Roos BA, Ong ES, Evans RM (March 1982). "Calcitonin mRNA polymorphism: peptide switching associated with alternative RNA splicing events". Proceedings of the National Academy of Sciences of the United States of America. 79 (6): 1717–21. Bibcode:1982PNAS...79.1717R. doi:10.1073/pnas.79.6.1717. PMC 346051. PMID 6952224. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Schmucker D, Clemens JC, Shu H, Worby CA, Xiao J, Muda M, Dixon JE, Zipursky SL (June 2000). "Drosophila Dscam is an axon guidance receptor exhibiting extraordinary molecular diversity". Cell. 101 (6): 671–84. doi:10.1016/S0092-8674(00)80878-8. PMID 10892653. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Matlin AJ, Clark F, Smith CW (May 2005). "Understanding alternative splicing: towards a cellular code". Nature Reviews. Molecular Cell Biology. 6 (5): 386–98. doi:10.1038/nrm1645. PMID 15956978. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Sammeth M, Foissac S, Guigó R (August 2008). Brent MR (المحرر). "A general definition and nomenclature for alternative splicing events". PLoS Computational Biology. 4 (8): e1000147. Bibcode:2008PLSCB...4E0147S. doi:10.1371/journal.pcbi.1000147. PMC 2467475. PMID 18688268. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Clark D (2005). Molecular biology. Amsterdam: Elsevier Academic Press. ISBN 978-0-12-175551-5. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Reid DC, Chang BL, Gunderson SI, Alpert L, Thompson WA, Fairbrother WG (December 2009). "Next-generation SELEX identifies sequence and structural determinants of splicing factor binding in human pre-mRNA sequence". RNA. 15 (12): 2385–97. doi:10.1261/rna.1821809. PMC 2779669. PMID 19861426. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wang Z, Burge CB (May 2008). "Splicing regulation: from a parts list of regulatory elements to an integrated splicing code" (Free full text). RNA. 14 (5): 802–13. doi:10.1261/rna.876308. PMC 2327353. PMID 18369186. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Barash Y, Calarco JA, Gao W, Pan Q, Wang X, Shai O, Blencowe BJ, Frey BJ (May 2010). "Deciphering the splicing code". Nature. 465 (7294): 53–9. Bibcode:2010Natur.465...53B. doi:10.1038/nature09000. PMID 20445623. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lyko F, Foret S, Kucharski R, Wolf S, Falckenhayn C, Maleszka R (November 2010). Keller L (المحرر). "The honey bee epigenomes: differential methylation of brain DNA in queens and workers". PLoS Biology. 8 (11): e1000506. doi:10.1371/journal.pbio.1000506. PMC 2970541. PMID 21072239. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Li-Byarlay H (2016). "The Function of DNA Methylation Marks in Social Insects". Frontiers in Ecology and Evolution. 4: 57. doi:10.3389/fevo.2016.00057. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية