مستقبل سطح الخلية
مستقبلات سطح الخلية (المستقبلات الغشائية، المستقبلات عبر الغشائية) هي مستقبلات مدمجة في الغشاء البلازمي للخلايا، وتعمل في تأشير الخلية عبر استقبال (الارتباط بـ) جزيئات خارج خلوية، وهي بروتينات غشائية مدمجة متخصصة تسمح بالاتصال بين الخلية والفضاء خارج الخلوي. يمكن أن تكون الجزيئات خارج الخلوية هرمونات، نواقل عصبية، سيتوكينات، عوامل نمو، جزيئات التصاق الخلية أو مغذيات. والتي تتفاعل مع المستقبل لتُحدث تغييرات في أيض ونشاط الخلية. في عملية توصيل الإشارة، يسبب ارتباط الربائط تسلسلا من التغيرات الكيميائية عبر الغشاء الخلوي.
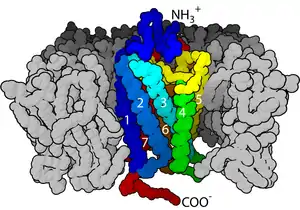
البنية والآلية
العديد من المستقبلات الغشائية هي بروتينات عبر غشائية، وتوجد أنواع عدة منها بما في ذلك البروتينات السكرية والبروتينات الليبيدية.[1] تم التعرف على المئات من المستقبلات ومازال العديد منها لم تتم دراستها والتعرف على وظائفها.[2][3] تصنف المستقبلات الغشائية عادة حسب بنيتها الثالثية (ثلاثية الأبعاد)، وإن كانت بنيتها الثالثية غير معروفة فيمكن أن تُصنف حسب تموضعها في الغشاء. في المستقبلات البسيطة، تعبر السلاسل الببتيدية الليبيد ثنائي الطبقة مرة واحدة، بينما يمكن أن تعبره في بروتينات أخرى مثل المستقبلات المقترنة بالبروتين ج حتى سبع مرات. يمكن أن يحتوي كل غشاء خلوي على عدة أنواع من المستقبلات الغشائية ذات انتشارات مختلفة على سطحه، كما يمكن لمستقبل واحد أن يكون منتشرا بشكل مختلف في عدة مناطق مختلفة من الغشاء وذلك حسب نوع الغشاء ووظيفة الخلية. غالبا ما تتكدس المستقبلات في منطقة على سطح الغشاء بدل أن تكون منتشرة بشكل متساوٍ.[4][5]
الآلية
تم اقتراح نموذجين لتفسير آلية عمل المستقبلات عبر الغشائية.
- الديمرة (المثنوية): يقترح نموذج الثتني أنه قبل ارتباط الربيطة، تتواجد المستقبلات على هيئة موحودية. وحين يحدث ارتباط الناهضة ترتبط الموحودات لتشكل مثنويا نشطا.
- الاستدارة: يُحدث ارتباط الربيطة بالجزء خارج الخلوي من المستقبل استدرة (تغيرا بنويا) في جزء من لوالب المستقبل عبر الغشائي.تغير هذه الاستدارة أجزاء المستقبل المشكوفة على الجانب داخل الخلوي من الغشاء، وهذا يغير طريقة تآثر المستقبل مع بروتينات أخرى داخل الخلية.[6]
مراجع
- Cuatrecasas P. (1974). "MEMBRANE RECEPTORS". Annual Review of Biochemistry. 43: 169–214. doi:10.1146/annurev.bi.43.070174.001125. PMID 4368906. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Dautzenberg FM, Hauger RL (February 2002). "The CRF peptide family and their receptors: yet more partners discovered". Trends Pharmacol. Sci. 23 (2): 71–7. doi:10.1016/S0165-6147(02)01946-6. PMID 11830263. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Rivière S, Challet L, Fluegge D, Spehr M, Rodriguez I (May 2009). "Formyl peptide receptor-like proteins are a novel family of vomeronasal chemosensors". نيتشر. 459 (7246): 574–7. doi:10.1038/nature08029. PMID 19387439. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Rothberg K.G.; Ying Y.S.; Kamen B.A.; Anderson R.G. (1990). "Cholesterol controls the clustering of the glycophospholipid-anchored membrane receptor for 5-methyltetrahydrofolate". The Journal of Cell Biology. 111 (6): 2931–2938. doi:10.1083/jcb.111.6.2931. PMC 2116385. PMID 2148564. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Jacobson C.; Côté P.D.; Rossi S.G.; Rotundo R.L.; Carbonetto S. (2001). "The Dystroglycan Complex Is Necessary for Stabilization of Acetylcholine Receptor Clusters at Neuromuscular Junctions and Formation of the Synaptic Basement Membrane". The Journal of Cell Biology. 152 (3): 435–450. doi:10.1083/jcb.152.3.435. PMC 2195998. PMID 11157973. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Maruyama, Ichiro N. (2015-09-01). "Activation of transmembrane cell-surface receptors via a common mechanism? The "rotation model"". BioEssays. 37 (9): 959–967. doi:10.1002/bies.201500041. ISSN 1521-1878. PMC 5054922. PMID 26241732. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة علوم عصبية
بوابة علوم عصبية بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية
 صور وملفات صوتية من كومنز
صور وملفات صوتية من كومنز