استدامة إنزيم اللاكتيز
استدامة إنزيم اللاكتيز هي النشاط المستمر لإنزيم اللاكتيز في فترة البلوغ . منذ ذلك الحين فإن الوظيفة الوحيدة لإنزيم اللاكتيز هي هضم سكر اللاكتوز الموجود في الحليب، في معظم أنواع الثدييات، يقل نشاط هذا الإنزيم بشكل كبير في فترة الفطام.[1] في بعض المجتمعات البشرية تطورت استدامة اللاكتيز كتلاؤم لاستهلاكهم [2] مصادر غير مصنعة من حليب الإنسان ومنتجات الألبان خلال فترة الطفولة.معظم المجتمعات في جميع أنحاء العالم بقوا من غيرالمستديمين لإنزيم اللاكتيز،[1] وكنتيجة لذلك يتأثرون بدرجات المتفاوتة بالحساسية المفرطة لسكر اللاكتوز عندما يصلون فترة البلوغ. مع ذلك ليس جميع الأشخاص الذين يعانون من عدم استدامة اللاكتيزبشكل وراثي يعانون من الحساسية المفرطة للاكتوز، وليس جميع أصحاب الحساسية المفرطة للاكتوز يحملون الطراز الجيني لعدم استدامة إنزيم اللاكتيز .
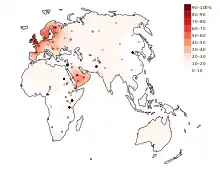
الانتشار العالمي للطراز الظاهري لعدم استدامة إنزيم اللاكتيز
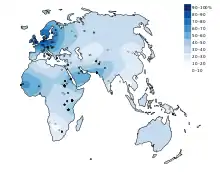
انتشار الطراز الظاهري لاستدامة اللاكتيز، أو القدرة على هضم سكر اللاكتوز في فترة البلوغ، ليست متجانسة في العالم . تكرار استدامة اللاكتيز في العالم متقلبة بشكل كبير . في أوروبا، انتشار الطراز الظاهري لاستدامة اللاكتيز يعتبر منحدرًا بتكرارات تتراوح من 15-54% في لجنوب الشرقي إلى 89-96% في الشمال الغربي،[3] على سبيل المثال، فقط 17% من اليونانيين، 14 % سردينيا يتوقع أن يحملون هذا الطراز، بينما 80 % من الفلبيين والهنغاريين و 100% من الإيرلنديين يتوقع أن يكونوا مستديمي اللاكتيز . تكرارات عالية من استدامة اللاكتيز أيضا تتواجد في بعض أماكن جنوب الصحراء الكبرى في إفريقيا والشرق الأوسط. لكن الوضع الأكثر شيوعا هو استدامة اللاكتيز المتوسط إلى المنخفضة: المتوسطة (11-32% ) في آسيا الوسطى ،[4] المنخفضة(أقل أو يساوي5%) في الهنود الحمر، الشرق الأسيوين، معظم المجتمعات الصينية وبعض المجتمعات الإفريقية .[3][4][5]
في إفريقيا انتشار استدامة اللاكتيز تعتبر غير منتظمة،[3][6][7] تقلبات كبيرة في التكرار تلاحظ في المجتمعات المتجاورة، مثلا بين بيجا ونيلوتس من السودان.[8] وهذا يجعل دراسة استدامة إنزيم اللاكتيز أكثر تعقيدًا .[5] نسب عالية من الطراز الظاهري لاستدامة اللاكتيز وجدت بين السكان الرعويين مثل شعب الفولاني والبدو .[3][9]
استدامة انزيم اللاكتيز تنتشر في شعب النغوي وبعض مجتمعات الرعاة في جنوب إفريقيا نظرا لاستهلاكهم منتجات الحليب في وجباتهم الغذائية . استدامة اللاكتيز بين أفراد نغوي أقل انتشارا بين مجتمعات شمال أوروبا لأن استهلاكهم التقليدي لمنتجات الحليب يكون على شكل أماسي(الحليب المخمّر- و يعرف بالماس بين الأفريقيين ) ويحتوي على نسبة أقل من اللاكتوز مقارنة بالحليب الطازج نتيجة لعملية التخمر التي يخضع لها .[9][10][10][11][12][13][14][15][16][17]
علم الوراثة
أظهرت دراسات عديدة أن وجود طرازين ظاهريين، استدامة اللاكتيز ( الطراز المستمد )و عدم استدامة اللاكتيز ( نقص اللاكتيز ) يكون مبرمج جينيا. وأن استدامة اللاكتيز ليست بالضرورة متعلقة باستهلاك اللاكتوز بعد فترة الرضاع.[18][19]
طراز استدامة اللاكتيز تتضمن تعبيرا عاليا لجزيء mRNA، نشاطا عاليا للاكتيز، بالتالي القدرة على هضم اللاكتوز .بينما طراز عدم استدامة اللاكتيز يتضمن تعبيرا قليلا من جزيء mRNA ونشاطا أقل للاكتيز.[20] يتم تشفير إنزيم لاكتيز من الجين LCT.[18]
نقص اللاكتيز يتم توارثه عبر جينات جسمية متنحية وهذا يعني أن الأشخاص غير مستديمي اللاكتيز هم متماثلون جينيا يلزم وجود نسختين من جين النشاط المنخفض للاكتيز لظهور الصفة ( ألليل الأسلاف ),[18] من آبائهم الذين يكونون متماثلين أو غير متماثلين جينيا للصفة.[18][19] جين واحد فقط لنشاط -اللاكتيز المرتفع يلزم لظهور طراز استدامة اللاكتيز .[18][19] صفة استدامة اللاكتيز تعتبر صفة سائدة لأنه فقط نصف المستويات من نشاط اللاكتيز كافية لهضم اللاكتوز .المناطق غير المشفرة من DNA التي لا يتم نسخها هي المسؤولة عن طراز نقص اللاكتيز.أظهرت الدراسات وجود فقط ثماني حالات لأطفال مستديمي اللاكتيز من آباء يعانون من نقص اللاكتيز . بينما العديد من التنوع الجيني والعوامل الغذائية يحددان التعبير الجيني للاكتيز، لا يوجد أي دليل على وجود تغيرات تكيفية لإنتاج اللاكتيز في الفرد كإستجابة لتغير مستويات استهلاك اللاكتوز.[1] الطرازين الظاهريين المحددان لنقص اللاكتيز هما : الطراز الظاهري الأول، يمتاز بنقص في صنع مصدر LPH، والطراز الظاهري الثاني يرتبط بالصنع المفرط للمصدر، يصاحبه نقص في تحويل البروتين إلى شكله الجزيئي الناضج .[21] يمتلك إنزيم اللاكتيز موقعين نشطين لتحليل سكر اللاكتوز، الأول على Glu1273 والثاني على Glu1749، اللذان يعملان بشكل منفصل على تكسير سكر اللاكتوز إلى نوعين منفصلين من الجزيئات .[1]
على الأقل ست طفرات ( طفرة استبدال نيوكليوتيد ) ترتبط بالتعبير الجيني لإنتاج إنزيم اللاكتيز.[22] تقع جميعها على الجين في المنطقة MCM6 المضادة ل جين اللاكتيز (LCT). تعتبر هذه المنطقة منطقة محفزة لنسخ جين اللاكتيز.[23][24][25] أول بديل جيني مرتبط باستدامة اللاكتيز تم تحديده هو C/T*−13910.[26] جين الأسلاف هو C والمستمد المرتبط باستدامة اللاكتيز هو T. في نفس الدراسة تم تحديد بديل جيني آخر يرتبط أيضا بالطراز الظاهري للصفة في الكثير من الحالات هو : G*/A-22018.
تم تحديد أليلات أخرى مرتبطة باستدامة اللاكتيز : G/C*-14010,C/G*-13907 [9] و T/G*-13915.[7][9][27] هذا الأليل يوصف بأنه جزء من الأليل المركب T/C*3712.[28] هذه البدائل الثلاثة تنتشر بشكل واسع في بعض المجتمعات.[29] البدائل النادرة تم تحديدها في دراسات قليلة، مثل G/A*14107 في الاكسوزا[22] والفولاني من مالي .[22]
أليلات استدامة اللاكتيز تختلف في توزيعها الجغرافي . في أوروبا ومجتمعات الأسلاف الأوروبيين تكون مرتبطة بالطفرة −13,910 C/T في المنطقة المحفزة لتشفير جين اللاكتيز. هذا يختلف من انتشار أليلات استدامة اللاكتيز في بقية أنحاء العالم .خصوصا في إفريقيا والشرق الأوسط، حيث يظهر العديد من الأليلات .
توجد أليلات T/G*-13915 في مجتمعات شرق وشمال إفريقيا[30] والشرق الأوسط . تم تحديد أليل G/C*-14010 في شرق إفريقيا . الأليل C/G*13907 تم وصفه في السودان واثيوبيا.[9][27][31] الأليل الأوروبي تم تحديده أيضا في بعض مجتمعات إفريقيا مثل مجتمعات الفولاني من مالي،[22] السودان[32] والكاميرون.[27] وجنوب إفريقيا،[14][16] تم تحديد هذا الأليل أيضا في آسيا الوسطى .[4]
لم يتم تحديد كيف تعمل البدائل السابق ذكرها في تنظيم التعبير الجيني لإنزيم اللاكتيز بالضبط . لم يتم تحديد أي من الطفرات حصريا لظاهرة استدامة اللاكتيز، ومن المحتمل أنه هناك العديد من الأليلات من الممكن اكتشافها .[33] لو ركزنا على ( البديل الأوروبي ) الموقع −13910 له وظيفة تحفيزية لمنظم اللاكتيز ( المنظم يسهل نسخ جين اللاكتيز ). T−13910 يعتبر محفزا أكبر من C−13910، لذا تعتبر هذه الطفرة هي المسؤولة عن التعبيرات الجينية المختلفة لإنزيم اللاكتيز،[34] بالرغم من أنه ليس دليلا كافيا يثبت أن استدامة اللاكتيز تنتج فقط بوجود C−13910→T−13910.[18]
بالإضافة، أظهرت دراسة أجريت على المجتمع الفنلندي أن جين اللاكتيز له تعبيرا جينيا أكبر عندما يندمج G−22018 مع T-13910.[18]
الفوائد التطورية
استدامة اللاكتيز هي مثال على الاختيار الطبيعي في البشر، تم تسجيله على أنه أقوى عامل اختيار من أي جين بشري آخر .[18] الأسباب الخاصة التي تضفي الأهمية الاختيارية لاستدامة اللاكتيز ما زالت مطروحة للتفكر .[35]
وضعت أدلة عديدة على الاختيار الإيجابي المؤثر على الأليل T*-13910، إنه يقع على التماثل الجيني c. 1 Mb.قوة الاختيار مماثلة لتلك المقدرة لمقاومة الملاريا . تم إنشاء الاستدلالات الفردية بناء على المعلومات من مجتمعات آسيا الوسطى، حيث تم تحديد الاختيار هناك جيدا، على أية حال فهو أقل قوة من المجتمعات الأوروربية. بالتالي، لو أن T*13910 ليس هو المسبب لاستدامة اللاكتيز، فإنه سيتم اختياره خلال التاريخ التطوري البشري. أثبت أن البدائل الأخرى خضعت أيضا للاختيار . يقع الأليل C*-14010 على امتداد القطعة المتماثلة جينيا (> 2 Mb). الأليل المركب G*-13915 & C*-3712 ثبت أنه يقع على امتداد القطعة المتماثلة جينيا (1.1 Mb[10] to 1.3 Mb[28]).
القابلية لهضم اللاكتوز ليست بدعة بشرية، جميع الثدييات تقريبا تبدأ حياتها بقدرتها على هضم اللاكتوز. هذه الصفة ذات أهمية خلال مرحلة الرضاعة، لأن الحليب في هذه المرحلة هو الغذاء الرئيسي.عند ظهور الفطام، وتم إدخال الطعام إلى الوجبات الغذائية، قل استهلاك الحليب.نتيجة لذلك هضم اللاكتوز لم يعد ذو أهمية محددة.اتضح هذا عند اختبار الجين المنتج للاكتيز، الذي يقل تشفيره بعد فترة الفطام، مما ينتج عنه انخفاض في إنتاج إنزيم اللاكتيز.عندما يتم إنتاج هذه الإنزيمات بكميات قليلة، تظهر عدم استدامة اللاكتوز.
القدرة على هضم الحليب الطازج في مرحلة البلوغ تحدَد جينيا بواسطة البدائل المختلفة الواقعة أعلى جين اللاكتيز والتي تختلف بين المجتمعات، هذه البدائل توجد ضمن تكرارات عالية جدا في بعض المجتمعات وتظهر دليلا مقاما على الاختيار.هناك فرضيتان ملاحظتان بنظريات غير متشابهة والتي تحاول تفسير لماذا تم الاختيار الإيجابي للطرز الظاهرية لاستدامة اللاكتيز، الأولى تدعى الفرضية المجتمعية التاريخية، والتي تنص على أن السبب الرئيسي لاستدامة اللاكتيز هو إدخال مشتقات الحليب على المنتجات الغذائية. بينما الفرضية السببية المعاكسة تجادل على أن المجتمعات ذات التكرار العالي من استدامة اللاكتيز هي من تبنّت استهلاك مشتقات الحليب .
فرضية التطور المشترك للجينات
اعتمدت فرضية التطور المشترك للجينات للاختيار الإيجابي لاستدامة اللاكتيز على ملاحظة مجتمعات الرعاة الذين يظهرون مستويات عالية لاستدامة إنزيم اللاكتيز .بالرجوع لهذه الفرضية، فإن سبب الاختيار هو الميزة الغذائية لكونهم مستديمي للاكتيز .الأفراد الذين يظهرون الطراز الظاهري لاستدامة اللاكتيز يتمتعون بميزة حقيقية في الاكتساب الغذائي. وهذه الميزة خاصة بالمجتمعات التي يشكل فيها الحيوانات المنتجة للحليب والرعاة الحياة الأساسية. الدمج بين الرعي وجينات استدامة اللاكتيز تعطي الأفراد ميزة بناء المسكن، هذا يعني أنه سيقل التنافس على المصادر عن طريق اشتقاق مصادر غذائية ثانوية، الحليب. الحليب كمصدر غذائي ربما يكون ذو قيمة أكثر من اللحوم، فمعدل تجدده أسرع.بدلا من تربية وذبح الحيوانات، يمكن حفظ بقرة واحدة أو نعجة كمصدر بوقت وطاقة أقل .القيمة التنافسية التي يمنحها الأفراد الذين يتحملون اللاكتوز تزيد قوة اختيار الطراز الجيني لهذه الصفة، خاصة في أوقات الجفاف والمجاعة، وهذا بدوره يزيد إنتاج استدامة اللاكتيز بين أفراد المجتمع. على عكس المجتمعات التي لا تظهر أنشطة رعوية، فلا يوجد أي فائدة اختيارية لاستدامة اللاكتيز . الطفرات التي تنتج تنوعات أليلية والتي تنتج إنزيم اللاكتيز في فترة البلوغ هي ببساطة طفرات متعادلة.ولا تكسب أي ميزة تتعلق بلياقة الأفراد.نتيجة لذلك لا يوجد اختيار خلّد انتشار هذه التنوعات الجينية، وبقي الطرازان الظاهري والجيني لاستدامة اللاكتيز نادران.على سبيل المثال في شرق آسيا شهدت المصادر التاريخية أن الصينيين لا يستهلكون الحليب، بينما البدو الذين يعيشون في الضواحي يستهلكونه. هذا يعكس التوزيعات الحديثة للتعصب، يلاحظ أن الصين بلدا يفتقر للتسامح، بينما في مغنوليا والسهوب الآسيوية، الحليب ومشتقاته هو الغذا الرئيسي. يقوم البدو أيضا بصنع المشروبات الكحولية، التي تسمى إيراج أو كومس، من حليب الفرس، بالرغم من أن عملية التخمير تقلل نسبة سكر اللاكتوز الموجودة. يوجد سيناريوهان افترضا لفرضية التطور المشترك للجينات : إما أن استدامة اللاكتيز تطورت وتم اختيارها منذ بداية الممارسات الرعوية (الفرضية الثقافية- التاريخية) أو أن الرعي انتشر في المجتمعات التي استدامة اللاكتيز فيها أصلا مرتفعة ( فرضية السبب-العكسي). هناك استثناءات للفرضية مثل الصيادين التنزانيين الذين تنتشر بينهم استدامة اللاكتيز بنسبة 50%.
فوائد استدامة اللاكتيز في فترة البلوغ
استهلاك اللاكتوز له فائدة على استدامة اللاكتيزخلال فترة البلوغ، على سبيل المثال، في دراسة لقلب وصحة المرأة البريطانية ( 2009) بحثت الآثار في صحة الأليلات المشفرة لاستدامة اللاكتيز في المرأة، حيث الأليل C يظهر عدم استدامة اللاكتيز والأليل T ينتج استدامة اللاكتيز، وجدت الدراسة أن النساء المتماثلات جينيا للأليل C يمتلكون صحة أسوأ من النساء اللاتي يحملن الأليلان C و T أو نسختان من الأليل T.النساء اللاتي يحملن الطراز الجيني CC يمتلكون وركًا وكسورا في الزند، عظام أكثر هشاشة، أعتام أكبر في عدسة العين مقارنة بالمجموعات الأخرى. وهن تقريبا أقصر من بقية النساء بمعدل 4-6 مم، كما أنهن أقل وزنًا. بالإضافة إلى عوامل أخرى مثل العوامل الأيضية، المستوى الاجتماعي والاقتصادي، نمط الحياة، والخصوبة وجدت انها لا صلة لها بالنتائج، لذلك يمكن الاستنتاج أن استدامة اللاكتيز تعود بالنفع على اصحة النساء اللواتي يستهلكن منتجات الحليب ويمتلكون استدامة لإنزيم اللاكتيز.
فرضية امتصاص الكالسيوم
احتمال آخر هو فرضية امتصاص الكالسيوم.يفضل اللاكتوز الامتصاص المعوي للكالسيوم: فهو يساعد على تكوينه بشكله المذاب. هذا له ميزة في المناطق ذات الإشعاع الشمسي المنخفض حيث أن فيتامين د ضروري لتحويل الكالسيوم ويعتبر عاملا محددا. العلاقة بين استدامة اللاكتيز وخط العرض في 33 مجتمع سكاني في أوروبا وجدت مهمة وذات معنى، بينما لم تظهر هذه العلاقة بين استدامة اللاكتيز وخط الطول، وهذا يفترض أن امتصاص الكالسيوم ذو أهمية في المناطق التي يكون فيها الإشعاع الشمسي منخفضا في الشمال الأوروبي. ارتفاع امتصاص الكالسيوم يساعد في تقليل كساح الأطفال وترقق العظام.
فرضية المناخ الجاف
فرضت دراسة اختيار استدامة اللاكتيز بشكل مواز لمقاومة الملاريا في الفلوني من مالي .الآليات المفترضة : القيمة الغذائية للحليب، الكمية القليلة ل p- حمض الأمينوبنزويك مقارنة بالوجبات اللاحليبية، تناول الحليب المدعم مناعيا.
عدم استدامة اللاكتيز في الشعوب المعتمدة على الحليب
بالرغم من مناقشة ميزات الاختيار لاستدامة اللاكتيز، هناك دراسات لمجموعات عرقية التي تعتمد بشكل أساسي على استهلاك الحليب، تظهر تكرارات منخفضة لاستدامة اللاكتيز. في دراسة أجريب على 303 فرد من قبائل البيجا و 282 فرد من القبائل النيلية في السودان أظهرت وجود فرق حاد في انتشار الطراز الشكلي لاستدامة اللاكتيز في المجتمعين.تم تحديد استدامة اللاكتيز بواسطة اختبار نفس الهيدروجين. تكرار أيض اللاكتوز كان 18.4% في أعضاء قبائل البيجا لأعمار فوق الثلاثين، و 73.3% في القبائل النيلية لأعمار فوق الثلاثين.
التاريخ التطوري
بالرجوع إلى فرضية التطور المشترك للجينات، قابلية هضم اللاكتوز في البلوغ ( استدامة اللاكتيز ) أصبح ذو قيمة بعد ابتكار تربية الحيوانات وترويض الحيوانات المنتجة بشكل مستمر للحليب. مجتمعات الصيادين وقبل ثورة العصر الحجري أظهروا حساسية مفرطة للاكتوز وبأغلبية ساحقة، تماما كالصيادين الحديثين.أظهرت الدراسات الجينية أن الطفرات القديمة المرتبطة باستدامة اللاكتيز وصلت لمستويات يمكن تقديرها في المجتمعات البشرية في العشر آلاف سنة مضت، هذا يتوافق مع بداية ترويض الحيوانات الذي ظهر خلال ثورة العصر الحجري.لذلك تعتبر استدامة اللاكتيز كمثال على التطور البشري الحديث. واستدامة اللاكتيز كصفة جينية لكن صفة ثقافية في تربية الحيوانات، تم إنشاء فرضية التطور المشترك للجينات في العلاقة التعايشية بين الحيوان والإنسان مع تقدم الزراعة.
بالاعتماد على المجتمعات، إحدى الفرضيات أو الأخرى للاختيار الإيجابي لاستدامة اللاكتيز هي ذات صلة أكثر : في الشمال الأوروبي ربما تكون فرضية امتصاص الكالسيوم من العوامل التي تزيد قوة المعاملات الرياضية للاختيار، بينما في المجتمعات الإفريقية، حيث لا يعتبر نقص فيتامين د قضية، فإن انتشار الأليل يتوافق مع السعرات الحرارية المضافة والتغذية الرعوية.تم تحديد العديد من العلامات الجينية لاستدامة اللاكتيز، وهذه أظهرت أن لاستدامة اللاكتيز أصول متعددة في جميع أنحاء العالم ( هذا مثال على التطور التقاربي). على العموم، تم افتراض أن البديل T*13910 ظهر مرتين على الأقل بشكل مستقل .في الواقع لوحظ على فرضيتين مختلفتين : H98هو الأكثر شيوعا ( بين الآخرين في الفنلنديين والفلوني) و H8 H12 يرتبط بالمجتمعات المقيدة جغرافيا. النسخة المشتركة قديمة نسبيا. البديل H98 - الأكثر شيوعا بين الأوروبيين - قدّر ليرتفع إلى تكرارات معنوية 7,500 سنة مضت في وسط البلقان و أوروبا الوسطى ، الوقت والزمن مطابقان تماما لصناعة الفخار الأثرية . يوجد البديل T*13910 في الشمال الإفريقي أيضًا، لذا فإنه نشأ قبل 7500 في الشرق القريب، لكن المزارعون القدامى لم يمتلكوا مستويات عالية من استدامة اللاكتيز ولم يستهلكوا الحليب غير المعالج .بعض الفرضيات تعود للتاريخ التطوري لاستدامة اللاكتيز في بعض المناطق من العالم تم وصفها في الأسفل.
أوروبا
في أوروبا على وجه الاهتمام، النموذج الذي افترض لاستدامة اللاكتيز يجمع آليات الاختيار والآليات السكانية، بعض الدراسات استخدمت النمذجة لدراسة انسياق الجينات.بالاعتماد على بعض النماذج، انتشار استدامة اللاكتيز في أوروبا تعزى إلى الانسياق الجيني بشكل أساسي، الدلائل تيمكن أن تأتي أيضا من حقول أخرى، مثال على ذلك السجلات التاريخية : سجل المؤلفون الرومان أن سكان الشمال الأوروربي البريطان والألمان تحديدا شربوا الحليب غير المعالج، وهذا يتوافق تماما مع التوزيع الحديث لفرط الحساسية للاكتوز بين السكان في أوروربا، حيث أن السكان في ألمانيا وبريطانيا والدول الاسكندنافية أكثر تحملا للاكتوز، بينما هؤلاء الذين في جنوب أوروبا، إيطاليا تحديدا يعانون من عدم التحمل للاكتوز. انخفاض التحمل في جنوب أوروربا يمكن تفسيره عن طريق الانسياق الجيني فقط بينما التحمل الأعلى في شمال أوروبا يمكن أن يكون كنتيجة للاختيار الإيجابي. وجد المسح الجيني لظاهرة الاختيار وباستخدام DNA ل 230 من الغرب الآسيوي القديم والذين عاشوا بين 6500 و 300 قبل الميلاد أن الأليل المسؤول عن استدامة اللاكتيز ظهر في الأفراد الذين عاشوا وسط أوروبا في الفترة ما بين 2450 و 2140 قبل الميلاد.
آسيا الوسطى
في آسيا الوسطى، المسبب للتعدد الظاهري لاستدامة اللاكتيز مماثلة لتلك التي في أوروربا (T*13910, rs4988235)، على افتراض وجود الانتشار الجيني بين المنطقتين الجغرافيتين. وجد أن الأليل (T*13910) المسؤول عن استدامة اللاكتيز قد ازداد في آسيا الوسطى اعتمادا على انتشار استدامة اللاكتيز بتكرارات مرتفعة بين الكازاخ الذين يشكلون نسبة أقل من المجموع الجيني الغربي كما استدل من مجموع التحليل من القنوات الجسمية الدقيقة، هذا بدوره يمكن أن يكون دليلا جينيا غير مباشر على ترويض الخيول لإنتاج الحليب كما ورد في البقايا الأثرية.في الكازاخس الرعاة التقليديون قدرت استدامة اللاكتيز ب 25–32%، حيث أنه فقط 40.2% أظهروا الأعراض و 85–92% يحملون الأليل T*13910.
إفريقيا
الوضع في افريقيا أكثر تعقيدا ، حيث أن الخمس بدائل الرئيسية لاستدامة اللاكتيز وجدت.[9][13][14][36]
وجود أليلات T*13910 بين الرعاة يعزى إلى التدفق الجيني من أوروربا ، ووجود أليلات أخرى هو إشارات على التدفق الجيني من شرق إفريقيا.
تم افتراض أن البديل G*13915 انتشر من الشرق الأوسط ، بالارتباط مع ترويض الجمل العربي.
طفرة G-14009 أسست في أثيوبيا.
البديل G*13907 تركز بين متحدثي اللغة الافريوساتية في الشمال الشرقي الإفريقي.
C*14010 هو الأليل الأكثر شيوعا اليوم بين مجموعات الرعاة التي تحتل الشرق الأفريقي، المكان الذي نشروا الرعي فيه امتدادا لجنوب افريقيا .في النهاية ، أليل استدامة اللاكتيز (C*14010) يعتقد أنه وصل من الصحراء في المناطق التي قطن فيها متحدثو اللغة الافريوساتية.تم تتبع ظهور تربية الحيوانات والكلمات الدخيلة من اللغة الافريوساتية في لغات الصحراء النيلية ونجر-الكونغو . بالإضافة إلى وجود الدهون من الحليب المعالج على السيراميك في المواقع الأثرية الليبية ( توثيق بالكربون المشع c. 7,500 BP المتقارب لعمر طفرة C*14010). الطرق التطورية المسؤولة عن الانتشار السريع لاستدامة اللاكتيز مازالت غير معروفة بعد ، من بين السكان الذين يقطنون شرق افريقيا انتقلت استدامة اللاكتيز من ترددات مهملة إلى ترددات واسعة الانتشار في ثلاثة آلاف سنة فقط.على افتراض أن هذا ضغط اختياري قوي جدا . بعض الدراسات افترضت أيضا أن اختيار استدامة اللاكتيز ليس بهذه القوة المفترضة ( مسح اختياري ناعم)وأن قوته تتقلب حسب الظروف البيئية .
في الكائنات غير البشرية
يعتبر سوء امتصاص اللاكتوز نموذجيا في الثدييات البالغة، واستدامة اللاكتيز هي ظاهرة مرتبطة بإنتاج الألبان من قبل الإنسان.معظم الثدييات تفقد القدرة على هضم اللاكتوز عندما تكبر بالقدر الذي يمكنها من البحث عن غذائها بعيدا عن الأمهات.بعد الفطام أو الانتقال من مرحلة رضاعة الحليب إلى استهلاك الأغذية الأخرى ، تتناقص قدرتهم في الإنتاج الطبيعي لإنزيم اللاكتيز حيث تقل الحاجة له. على سبيل المثال، في الوقت الذي نما فيه الخنزير من 5 إلى 18 يوم في إحدى الدراسات ، فقد ما يقارب 67% من قابليته لامتصاص اللاكتوز، بينما يستطيع البشر هضم اللاكتوز طيلة الفترة الأولى لحياتهم الممتدة من 5-7سنوات.معظم الثدييات يتوقف إنتاج اللاكتيز عندها بوقت مبكر أكثر. تستطيع النعاج فطم نفسها عن حليب أمهاتها عندما تبلغ ستة أشهر إلى سنة من عمرها.الحمل تفطم نفسها بشكل منتظم عندما تبلغ 16 أسبوعا من عمرها.هذه الأمثلة تفترض أن ظاهرة استدامة اللاكتيز هي ظاهرة بشرية فريدة.
عوامل خارجية
وجدت بعض الأمثلة على عوامل مسؤولة عن استدامة اللاكتيز في غياب البديل الجيني المسؤول عن استدامة اللاكتيز.ربما يفتقر الأفراد للأليلات المسؤولة عن استدامة اللاكتيز، لكن يستطيعون تحمل اللاكتوز الموجود في المشتقات الحليبية التي يهضم فيها اللاكتوز بعملية التخمر ( كالجبنة واللبن)[37] بالإضافة إلى سلالة البكتيريا الصحية الموجودة في الأمعاء والتي تساهم في هضم اللاكتوز لتمكن الأفراد المفتقرون لجين استدامة اللاكتيز من الاستفادة من استهلاك الحليب.[37][38]
اختبار تحمل اللاكتوز
يجرى اختبار تحمل اللاكتوز من خلال الأسئلة، وفحص السريع لمستويات الغلوكوز خلال الليل وقبل الفحص، تعطى محاليل اللاكتوز في الصباح التالي للفحص كما يفحص مستوى غلوكوز الدم في فترة زمنية معينة. الناس الذين يظهرون زيادة حقيقية في مستويات الغلوكوز يعتبرون ذو قدرة على تحمل اللاكتوز.[16]
مراجع
- Swallow, Dallas M. (December 2003). "Genetics of Lactase Persistence and Lactose Intolerance". Annual Review of Genetics. 37 (1): 197–219. doi:10.1146/annurev.genet.37.110801.143820. PMID 14616060. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bersaglieri, Todd; Sabeti, Pardis C.; Patterson, Nick; Vanderploeg, Trisha; Schaffner, Steve F.; Drake, Jared A.; Rhodes, Matthew; Reich, David E.; Hirschhorn, Joel N. (June 2004). "Genetic Signatures of Strong Recent Positive Selection at the Lactase Gene". The American Journal of Human Genetics. 74 (6): 1111–1120. doi:10.1086/421051. PMC 1182075. PMID 15114531. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Gerbault, P.; Liebert, A.; Itan, Y.; Powell, A.; Currat, M.; Burger, J.; Swallow, D. M.; Thomas, M. G. (14 February 2011). "Evolution of lactase persistence: an example of human niche construction". Philosophical Transactions of the Royal Society B: Biological Sciences. 366 (1566): 863–877. doi:10.1098/rstb.2010.0268. PMC 3048992. PMID 21320900. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Heyer, Evelyne; Brazier, Lionel; Ségurel, Laure; Hegay, Tatiana; Austerlitz, Frédéric; Quintana-Murci, Lluis; Georges, Myriam; Pasquet, Patrick; Veuille, Michel (June 2011). "Lactase Persistence in Central Asia: Phenotype, Genotype, and Evolution". Human Biology. 83 (3): 379–392. doi:10.3378/027.083.0304. PMID 21740154. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Itan, Yuval; Jones, Bryony L; Ingram, Catherine JE; Swallow, Dallas M; Thomas, Mark G (2010). "A worldwide correlation of lactase persistence phenotype and genotypes". BMC Evolutionary Biology. 10 (1): 36. doi:10.1186/1471-2148-10-36. PMC 2834688. PMID 20144208. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Durham, William H. (1991). Coevolution: Genes, Culture, and Human Diversity. Stanford, CA: Stanford University Press. ISBN 9780804721561. مؤرشف من الأصل في 12 سبتمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ingram, Catherine J. E.; Mulcare, Charlotte A.; Itan, Yuval; Thomas, Mark G.; Swallow, Dallas M. (26 November 2008). "Lactose digestion and the evolutionary genetics of lactase persistence". Human Genetics. 124 (6): 579–591. doi:10.1007/s00439-008-0593-6. PMID 19034520. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bayoumi, R. A. L.; Flatz, S. D.; Kühnau, W.; Flatz, G. (June 1982). "Beja And Nilotes: Nomadic pastoralist groups in the Sudan with opposite distributions of the adult lactase phenotypes". American Journal of Physical Anthropology. 58 (2): 173–178. doi:10.1002/ajpa.1330580208. PMID 6810704. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Tishkoff, Sarah A; Reed, Floyd A; Ranciaro, Alessia; Voight, Benjamin F; Babbitt, Courtney C; Silverman, Jesse S; Powell, Kweli; Mortensen, Holly M; Hirbo, Jibril B; Osman, Maha; Ibrahim, Muntaser; Omar, Sabah A; Lema, Godfrey; Nyambo, Thomas B; Ghori, Jilur; Bumpstead, Suzannah; Pritchard, Jonathan K; Wray, Gregory A; Deloukas, Panos (10 December 2006). "Convergent adaptation of human lactase persistence in Africa and Europe". Nature Genetics. 39 (1): 31–40. doi:10.1038/ng1946. PMC 2672153. PMID 17159977. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Jones, Bryony Leigh; Oljira, Tamiru; Liebert, Anke; Zmarz, Pawel; Montalva, Nicolas; Tarekeyn, Ayele; Ekong, Rosemary; Thomas, Mark G.; Bekele, Endashaw; Bradman, Neil; Swallow, Dallas M. (9 June 2015). "Diversity of lactase persistence in African milk drinkers". Human Genetics. 134 (8): 917–925. doi:10.1007/s00439-015-1573-2. PMC 4495257. PMID 26054462. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Lactase persistence alleles reveal ancestry of southern African Khoe pastoralists". مؤرشف من الأصل في 5 مايو 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Origins of Lactase Persistence in Africa". مؤرشف من الأصل في 5 مايو 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Breton, Gwenna; Schlebusch, Carina M.; Lombard, Marlize; Sjödin, Per; Soodyall, Himla; Jakobsson, Mattias (April 2014). "Lactase Persistence Alleles Reveal Partial East African Ancestry of Southern African Khoe Pastoralists". Current Biology. 24 (8): 852–858. doi:10.1016/j.cub.2014.02.041. PMID 24704072. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Macholdt, Enrico; Lede, Vera; Barbieri, Chiara; Mpoloka, Sununguko W.; Chen, Hua; Slatkin, Montgomery; Pakendorf, Brigitte; Stoneking, Mark (April 2014). "Tracing Pastoralist Migrations to Southern Africa with Lactase Persistence Alleles". Current Biology. 24 (8): 875–879. doi:10.1016/j.cub.2014.03.027. PMC 5102062. PMID 24704073. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Africans' Ability to Digest Milk Co-Evolved with Livestock Domestication". مؤرشف من الأصل في 25 مايو 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ranciaro, Alessia; Campbell, Michael C.; Hirbo, Jibril B.; Ko, Wen-Ya; Froment, Alain; Anagnostou, Paolo; Kotze, Maritha J.; Ibrahim, Muntaser; Nyambo, Thomas; Omar, Sabah A.; Tishkoff, Sarah A. (April 2014). "Genetic Origins of Lactase Persistence and the Spread of Pastoralism in Africa". The American Journal of Human Genetics. 94 (4): 496–510. doi:10.1016/j.ajhg.2014.02.009. PMC 3980415. PMID 24630847. الوسيط
|CitationClass=تم تجاهله (مساعدة) - "Why 65% of Humans Can't Drink Milk". مؤرشف من الأصل في 5 مايو 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Troelsen JT (May 2005). "Adult-type hypolactasia and regulation of lactase expression". Biochim. Biophys. Acta. 1723 (1–3): 19–32. doi:10.1016/j.bbagen.2005.02.003. PMID 15777735. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Wang Y, Harvey CB, Hollox EJ, Phillips AD, Poulter M, Clay P, Walker-Smith JA, Swallow DM (June 1998). "The genetically programmed down-regulation of lactase in children". Gastroenterology. 114 (6): 1230–6. doi:10.1016/S0016-5085(98)70429-9. PMID 9609760. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Harvey CB, Wang Y, Hughes LA, Swallow DM, Thurrell WP, Sams VR, Barton R, Lanzon-Miller S, Sarner M (January 1995). "Studies on the expression of intestinal lactase in different individuals". Gut. 36 (1): 28–33. doi:10.1136/gut.36.1.28. PMC 1382348. PMID 7890232. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lloyd M, Mevissen G, Fischer M, Olsen W, Goodspeed D, Genini M, Boll W, Semenza G, Mantei N (February 1992). "Regulation of intestinal lactase in adult hypolactasia". J. Clin. Invest. 89 (2): 524–9. doi:10.1172/JCI115616. PMC 442883. PMID 1737843. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lokki, A Inkeri; Järvelä, Irma; Israelsson, Elisabeth; Maiga, Bakary; Troye-Blomberg, Marita; Dolo, Amagana; Doumbo, Ogobara K; Meri, Seppo; Holmberg, Ville (14 January 2011). "Lactase persistence genotypes and malaria susceptibility in Fulani of Mali". Malaria Journal. 10 (1): 9. doi:10.1186/1475-2875-10-9. PMC 3031279. PMID 21235777. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Lewinsky, Rikke H.; Jensen, Tine G.K.; Møller, Jette; Stensballe, Allan; Olsen, Jørgen; Troelsen, Jesper T. (15 December 2005). "T −13910 DNA variant associated with lactase persistence interacts with Oct-1 and stimulates lactase promoter activity in vitro". Human Molecular Genetics. 14 (24): 3945–3953. doi:10.1093/hmg/ddi418. PMID 16301215. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Jensen, Tine G. K.; Liebert, Anke; Lewinsky, Rikke; Swallow, Dallas M.; Olsen, Jørgen; Troelsen, Jesper T. (15 February 2011). "The −14010*C variant associated with lactase persistence is located between an Oct-1 and HNF1α binding site and increases lactase promoter activity". Human Genetics. 130 (4): 483–493. doi:10.1007/s00439-011-0966-0. PMID 21327791. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Enattah, Nabil Sabri; Jensen, Tine G.K.; Nielsen, Mette; Lewinski, Rikke; Kuokkanen, Mikko; Rasinpera, Heli; El-Shanti, Hatem; Seo, Jeong Kee; Alifrangis, Michael; Khalil, Insaf F.; Natah, Abdrazak; Ali, Ahmed; Natah, Sirajedin; Comas, David; Mehdi, S. Qasim; Groop, Leif; Vestergaard, Else Marie; Imtiaz, Faiqa; Rashed, Mohamed S.; Meyer, Brian; Troelsen, Jesper; Peltonen, Leena (January 2008). "Independent Introduction of Two Lactase-Persistence Alleles into Human Populations Reflects Different History of Adaptation to Milk Culture". The American Journal of Human Genetics. 82 (1): 57–72. doi:10.1016/j.ajhg.2007.09.012. PMC 2253962. PMID 18179885. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Enattah, Nabil Sabri; Sahi, Timo; Savilahti, Erkki; Terwilliger, Joseph D.; Peltonen, Leena; Järvelä, Irma (14 January 2002). "Identification of a variant associated with adult-type hypolactasia". Nature Genetics. 30 (2): 233–237. doi:10.1038/ng826. PMID 11788828. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ingram, Catherine J. E.; Elamin, Mohamed F.; Mulcare, Charlotte A.; Weale, Michael E.; Tarekegn, Ayele; Raga, Tamiru Oljira; Bekele, Endashaw; Elamin, Farouk M.; Thomas, Mark G.; Bradman, Neil; Swallow, Dallas M. (21 November 2006). "A novel polymorphism associated with lactose tolerance in Africa: multiple causes for lactase persistence?". Human Genetics. 120 (6): 779–788. doi:10.1007/s00439-006-0291-1. PMID 17120047. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Imtiaz, F; Savilahti, E; Sarnesto, A; Trabzuni, D; Al-Kahtani, K; Kagevi, I; Rashed, M S; Meyer, B F; Jarvela, I (1 October 2007). "The T/G 13915 variant upstream of the lactase gene (LCT) is the founder allele of lactase persistence in an urban Saudi population". Journal of Medical Genetics. 44 (10): e89. doi:10.1136/jmg.2007.051631. PMC 2597971. PMID 17911653. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Torniainen, Suvi; Parker, M Iqbal; Holmberg, Ville; Lahtela, Elisa; Dandara, Collet; Jarvela, Irma (2009). "Screening of variants for lactase persistence/non-persistence in populations from South Africa and Ghana". BMC Genetics. 10 (1): 31. doi:10.1186/1471-2156-10-31. PMC 2713916. PMID 19575818. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Gerbault, Pascale; Moret, Céline; Currat, Mathias; Sanchez-Mazas, Alicia; O'Rourke, Dennis (24 July 2009). "Impact of Selection and Demography on the Diffusion of Lactase Persistence". PLoS ONE. 4 (7): e6369. Bibcode:2009PLoSO...4.6369G. doi:10.1371/journal.pone.0006369. PMC 2711333. PMID 19629189. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Ingram, Catherine J. E.; Raga, Tamiru Oljira; Tarekegn, Ayele; Browning, Sarah L.; Elamin, Mohamed F.; Bekele, Endashaw; Thomas, Mark G.; Weale, Michael E.; Bradman, Neil; Swallow, Dallas M. (24 November 2009). "Multiple Rare Variants as a Cause of a Common Phenotype: Several Different Lactase Persistence Associated Alleles in a Single Ethnic Group". Journal of Molecular Evolution. 69 (6): 579–588. Bibcode:2009JMolE..69..579I. doi:10.1007/s00239-009-9301-y. PMID 19937006. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Enattah, Nabil Sabri; Trudeau, Aimee; Pimenoff, Ville; Maiuri, Luigi; Auricchio, Salvatore; Greco, Luigi; Rossi, Mauro; Lentze, Michael; Seo, J.K.; Rahgozar, Soheila; Khalil, Insaf; Alifrangis, Michael; Natah, Sirajedin; Groop, Leif; Shaat, Nael; Kozlov, Andrew; Verschubskaya, Galina; Comas, David; Bulayeva, Kazima; Mehdi, S. Qasim; Terwilliger, Joseph D.; Sahi, Timo; Savilahti, Erkki; Perola, Markus; Sajantila, Antti; Järvelä, Irma; Peltonen, Leena (September 2007). "Evidence of Still-Ongoing Convergence Evolution of the Lactase Persistence T-13910 Alleles in Humans". The American Journal of Human Genetics. 81 (3): 615–625. doi:10.1086/520705. PMC 1950831. PMID 17701907. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Itan Y.; Jones B. L.; Ingram C. J. E.; Swallow D. M.; Thomas M. G. (2010). "A worldwide correlation of lactase persistence phenotype and genotypes". BMC Evolutionary Biology. 10 (1): 36. doi:10.1186/1471-2148-10-36. PMC 2834688. PMID 20144208. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Troelsen JT, Olsen J, Møller J, Sjöström H (December 2003). "An upstream polymorphism associated with lactase persistence has increased enhancer activity". Gastroenterology. 125 (6): 1686–94. doi:10.1053/j.gastro.2003.09.031. PMID 14724821. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Itan, Yuval; Powell, Adam; Beaumont, Mark A.; Burger, Joachim; Thomas, Mark G.; Tanaka, Mark M. (28 August 2009). "The Origins of Lactase Persistence in Europe". PLoS Computational Biology. 5 (8): e1000491. Bibcode:2009PLSCB...5E0491I. doi:10.1371/journal.pcbi.1000491. PMC 2722739. PMID 19714206. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Liebert, A. (2014). Evolutionary and molecular genetics of regulatory alleles responsible for lactase persistence (PhD Thesis). University College London. مؤرشف من الأصل في 5 مايو 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Silanikove N, Leitner G, Merin U (2015). "The Interrelationships between Lactose Intolerance and the Modern Dairy Industry: Global Perspectives in Evolutional and Historical Backgrounds". Nutrients (Review). 7 (9): 7312–31. doi:10.3390/nu7095340. PMC 4586535. PMID 26404364. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Szilagyi, Andrew (13 August 2015). "Adaptation to Lactose in Lactase Non Persistent People: Effects on Intolerance and the Relationship between Dairy Food Consumption and Evalution of Diseases". Nutrients. 7 (8): 6751–6779. doi:10.3390/nu7085309. PMC 4555148. PMID 26287234. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة طب
بوابة طب
