إخصاب مزدوج
التخصيب المزدوج(بالانجليزية:Double fertilization) هو آلية معقدة إخصاب للنباتات المزهرة (كاسيات البذور. هذه العملية تشمل الانضمام إلى أنثى مشيج (megagametophyte ، وتسمى أيضا كيس الجنين) مع اثنين من الأمشاج الذكور (الحيوانات المنوية). يبدأ عندما تلتزم حبوب اللقاح بوصمة الكارب ، وهي البنية التناسلية الأنثوية للزهرة. ثم تأخذ حبوب اللقاح في الرطوبة وتبدأ تنبت ، وتشكل لقاح الأنبوب الذي يمتد نحو المبيض (نباتات) | مبيض) من خلال الأسلوب. ثم يدخل طرف أنبوب اللقاح المبيض ويخترق فتحة الميكروبية في البويضة. يستمر أنبوب حبوب اللقاح لتحرير الحيوانتين المنويتين في الفطور megagametophyte.
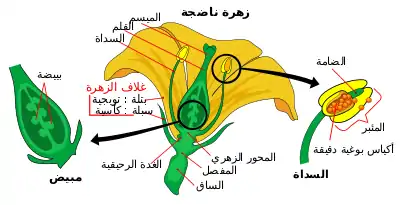

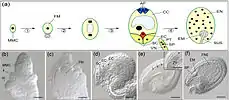
البويضات هي 8 في عدد ورتبت في شكل 3 + 2 + 3 (من أعلى إلى أسفل) أي 2 التآزر و 1 خلية بويضة، 2 الخلايا المركزية القطبية، 3 الخلايا المضادة للالتماس. تقوم إحدى الحيوانات المنوية بتخصيب خلية البويضة وتجمع الحيوانات المنوية الأخرى مع النقطتين القطبيتين للخلية المركزية 'للميغاجاميتوفيت. تجمع الحيوانات المنوية أحادية الصبغيات مع البيضة أحادية الصبغيات لتكوّن زايغوت ثنائي الصيغة الصبغية ، وهي العملية التي يطلق عليها تخصيب ، في حين أن الحيوانات المنوية الأخرى والنوعين أحادي الصبغة القطبية للخلية المركزية الكبيرة لشكل الغدة الكظرية المتوسطة (megagametophyte) نواة تصلب ثلاثي ( 'الاندماج الثلاثي' ). بعض النباتات قد تشكل نوى تعدد الصيغ الصبغية. ثم تتطور الخلية الكبيرة من المشيج إلى السويداء السويداء ، وهو نسيج غني بالمغذيات يوفر التغذية للجنين النامي. يتطور المبيض، المحيط بالبويضات، إلى ثمرة، التي تحمي البذور وقد تعمل لتفريقها.[1]
تنشأ النويتان الخلويتان المركزيتان للخلية (النواة القطبية) التي تساهم في الإندوسبيرم، عن طريق الانقسام الفتيلي من نفس المنتج ميوزيك الذي أدى إلى ظهور البويضة. مساهمة الأمومة في الدستور الجيني للاندوسبيرم ثلاثي الجذور هو ضعف مساهمة الجنين.
في دراسة أجريت في عام 2008 من النبات رشاد أذن الفأر '، تم توثيق هجرة النوى الذكرية داخل الأمشاج الأنثوية، في الاندماج مع الأنوية الأنثوية، لأول مرة باستخدام في الجسم الحي التصوير. كما تم تحديد بعض الجينات المشاركة في عملية الهجرة والدمج.[2]
وقد تم الإبلاغ عن دليل الإخصاب المزدوج في جنتوم ، وهي نباتات غير مزهرة.[3]
نبذة تاريخية موجزة
تم اكتشاف الإخصاب المزدوج منذ أكثر من قرن من قبل سيرجي نواسين وغريغنارد في كييف,[4] الإمبراطورية الروسية ، وليون غينغر في فرنسا. كل من صنع هذا الاكتشاف بشكل مستقل عن الآخر. [5] زنبق بنفسجي و بوقية tenella 'استخدمت في الملاحظات الأولى للتخصيب المزدوج، والتي تم صنعها باستخدام الكلاسيكية المجهر الضوئي . بسبب قيود المجهر الضوئي، كان هناك العديد من الأسئلة التي لم يتم الرد عليها فيما يتعلق بعملية الإخصاب المزدوج. ومع ذلك، مع تطوير المجهر الإلكتروني ، تمت الإجابة على العديد من الأسئلة. وأبرزها، أن الملاحظات التي أبدتها مجموعة و. جنسن أظهرت أن الأمشاج الذكور لم يكن لديهم أي جدران خلوية وأن غشاء البلازما في الأمشاج قريب من غشاء البلازما للخلية المحيطة به داخل حبوب اللقاح.[6]
تسميد مزدوج في Gymnosperms
هناك شكل بدائي أكثر بكثير من الإخصاب المزدوج يحدث في التكاثر الجنسي لأمر من عاريات البذور المعروفة باسم Gnetales.[3] على وجه التحديد، تم توثيق هذا الحدث في كل من الإيفيدرا و جنتوم '، وهي مجموعة فرعية من شعبة الجنتويات.[7] في افيدرا nevadensis ، '، يتم ترسب خلية منوية واحدة binucleate في خلية البويضة. بعد حدث التخصيب الأولي، يتم تحويل نواة الحيوانات المنوية الثانية لتخصيب نواة بيضة إضافية موجودة في سيتوبلازما البيض. في معظم نباتات البذور الأخرى، نجد أن "نواة القناة البطنية" الثانية عادة ما تكون عديمة الفائدة من الناحية الوظيفية.[8] في Gnetum gnemon ، توجد العديد من نوى البويضة الحرة في السيتوبلازم الأنثى داخل الأنثى نابتة عرسية. ونجحت في اختراق الاختام الأنثوي الناضج بواسطة [أنبوب اللقاح] ، السيتوبلازم الأنثوي والنواة الحرة للتحرك لإحاطة أنبوب اللقاح. يُطلق من الخلية المنوية ثنائية النواة نوعين من نواة الحيوانات المنوية التي تنضم بعد ذلك إلى نواة بيضة حرة لإنتاج زغيليتي حيويين قابلين للحياة، وهي خاصية متماثلة بين العائلات "الإيفيدرا" و "الجنيوم". [9] في كلا العائلتين، ينتج حدث التخصيب الثاني مضاعفا إضافيا الجنين. يتم إحباط هذا الجنين الزائد في وقت لاحق، مما يؤدي إلى توليف جنين واحد ناضجة فقط.[10] إن منتج الإخصاب الإضافي في "الإيفيدرا" لا يغذي الجنين الأساسي، حيث أن المشيمة الأنثوية مسؤولة عن توفير المغذيات.[9] تؤدي العملية الأكثر بدائية من الإخصاب المزدوج في جمنوسبيرم إلى نواتين ثنائيي الميل المغلق في نفس خلية البويضة. هذا يختلف عن حالة قثطرة نباتية ، مما يؤدي إلى فصل خلية البيض و السويداء.[11] بحوث جزيئية مقارنة حول جينوم "G. وقد كشفت gnemon أن gnetophytes ترتبط ارتباطا وثيقا الصنوبريات من أنها إلى كاسيات البذور.[12][13][14] إن رفض فرضية anthophyte ، التي تحدد gnetales و angiosperms هي الأخت taxa ، يؤدي إلى التكهنات بأن عملية الإخصاب المزدوج هو نتاج تطور متقارب ونشأ بشكل مستقل بين gnetophytes و angiosperms [15]
في المختبر مزدوجة الإخصاب
في كثير من الأحيان يتم استخدام الاخصاب المزدوج في المختبر لدراسة التفاعلات الجزيئية وكذلك الجوانب الأخرى لاندماج الأمشاج في النباتات المزهرة. واحدة من العقبات الرئيسية في تطوير التسميد المزدوج في المختبر بين الأمشاج الذكور والإناث هو حبس الحيوانات المنوية في أنبوب حبوب اللقاح والبيضة في كيس الجنين. وقد تم بالفعل تحقيق انصهار يتم التحكم فيه من البويضة والحيوانات المنوية مع نباتات الخشخاش.[16] إنبات حبوب اللقاح ، إدخال أنبوب اللقاح، وعمليات إخصاب مزدوجة تمت ملاحظتها كلها للمضي قدما بشكل طبيعي. في الواقع، لقد تم استخدام هذه التقنية بالفعل للحصول على البذور في العديد من النباتات المزهرة وكانت تسمى "إخصاب أنبوب الاختبار".[17]
الهياكل والوظائف ذات الصلة
Megagametophyte
تُدعى الأنثى المشيميت ، و megagametophyte ، التي تشارك في التخصيب المزدوج في كاسيات قزحية التي هي أحادية العدد الكروموسوم الجنين. يحدث هذا داخل البويضة ، يحيط بها المبيض عند قاعدة الكارب. يُحيط بـ megagametophyte (واحد أو اثنين) من العناصر المدمجة، والتي تشكل فتحة تسمى micropyle. ينشأ الفطر megagametophyte ، وهو عادة أحادي الصبغة الفردانية ، عن الخلية الأم (عادة صيغة صبغية megacore ، والتي تسمى أيضًا بالميغوبوروتية megasporocyte. يختلف التسلسل التالي للأحداث، حسب نوع معين، ولكن في معظم الأنواع، تحدث الأحداث التالية. وتخضع الخلية الكبرى لانقسام خلوي ناتج، ينتج أربعة أحواض سائبة أحادية الصيغة الصبغية. واحد فقط من أربعة أبطال ناجحين. يخضع هذا الميكوسور لثلاث جولات من الانقسام الانقسامي، مما يؤدي إلى سبع خلايا تحتوي على ثمانية نواة أحادية الصيغة الصبغية (تحتوي الخلية المركزية على نواتين تدعى النويات القطبية). يتكون الجزء السفلي من كيس الأجنة من خلية بويضة أحادية الصيغة الصبغية موضوعة في وسط خليتين أحاديتين مختلفتين، تدعى التآزرات. تعمل التآزرات في جذب وتوجيه أنبوب اللقاح إلى الميغاميتوفيت من خلال الميكروب. في الطرف العلوي من megagametophyte هي ثلاث خلايا مضادة للظواهر.
عرسية نباتية مكروية
وتوجد الذبابة الشبكية الذكرية، أو الفئران المصغرة (microgametophytes) ، التي تشارك في التخصيب المزدوج، داخل حبوب حبوب اللقاح. أنها تتطور داخل microsporangia ، أو حبوب اللقاح، من anthers على الأسدية. يحتوي كل ميكروسبورانجيوم على خلايا أم مضاعفة [ميكروسبريد] ، أو خلايا صغرى. تخضع كل خلية صغرى إلى الانقسام الاختزالي، لتشكل أربعة مجهرية أحادية الصبغيات، كل منها يمكن أن يتطور في النهاية إلى حبوب لقاح. يخضع microspore انقسام متساو و انقسام السيتوبلازم لإنتاج خليتين منفصلتين، الخلية المولدة والخلية الأنبوبية. هذان الخليتان بالإضافة إلى جدار بوغ يشكلان حبوب حبوب اللقاح غير الناضجة. عندما ينضج المشيج الذكري، تمر الخلية المولدة في الخلية الأنبوبية، وتخضع الخلية المولدة للانقسام، وتنتج خليتين من الحيوانات المنوية. وبمجرد أن تنضج حبوب اللقاح، ينفتح سداة ، ليطلق اللقاح. يتم حمل حبوب اللقاح إلى مدقة من زهرة أخرى، بواسطة الرياح أو ملقحات الحيوانات، وتوضع على الوصمة. عندما تنبت حبة اللقاح، تنتج خلية الأنبوب أنبوب حبوب اللقاح، الذي يطيل ويمتد إلى النمط الطويل من الكربل وإلى المبيض، حيث يتم إطلاق خلايا الحيوانات المنوية في الطبقة المتوسطة العملاقة. مضاعفات التسميد مزدوجة من هنا.[18]
المصادر
- Berger, F. (January 2008). "Double-fertilization, from myths to reality". Sexual Plant Reproduction. 21 (1): 3–5. doi:10.1007/s00497-007-0066-4. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Berger, F.; Hamamura, Y. & Ingouff, M. & Higashiyama, T. (August 2008). "Double fertilization – Caught In The Act". Trends in Plant Science. 13 (8): 437–443. doi:10.1016/j.tplants.2008.05.011. PMID 18650119. الوسيط
|CitationClass=تم تجاهله (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) -
V. Raghavan (September 2003). "Some reflections on double fertilization, from its discovery to the present". New Phytologist. 159 (3): 565–583. doi:10.1046/j.1469-8137.2003.00846.x. مؤرشف من الأصل في 13 يناير 2017. اطلع عليه بتاريخ 27 أغسطس 2013. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Kordium EL (2008). "[Double fertilization in flowering plants: 1898-2008]". Tsitol. Genet. (باللغة الروسية). 42 (3): 12–26. PMID 18822860. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Jensen, W. A. (February 1998). "Double Fertilization: A Personal View". Sexual Plant Reproduction. 11 (1): 1–5. doi:10.1007/s004970050113. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Dumas, C. & Rogowsky, P. (August 2008). "Fertilization and Early Seed Formation". Comptes Rendus Biologies. 331 (10): 715–725. doi:10.1016/j.crvi.2008.07.013. PMID 18926485. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Carmichael, J. S.; Friedman, W. E. (1995-12-01). "Double Fertilization in Gnetum gnemon: The Relationship between the Cell Cycle and Sexual Reproduction". The Plant Cell (باللغة الإنجليزية). 7 (12): 1975–1988. doi:10.1105/tpc.7.12.1975. ISSN 1040-4651. PMC 161055. PMID 12242365. مؤرشف من الأصل في 20 يونيو 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Friedman, William E. (1990). "Sexual Reproduction in Ephedra nevadensis (Ephedraceae): Further Evidence of Double Fertilization in a Nonflowering Seed Plant". American Journal of Botany. 77 (12): 1582–1598. doi:10.2307/2444491. JSTOR 2444491. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Carmichael, Jeffrey S.; Friedman, William E. (1996). "Double Fertilization in Gnetum gnemon (Gnetaceae): Its Bearing on the Evolution of Sexual Reproduction within the Gnetales and the Anthophyte Clade". American Journal of Botany. 83 (6): 767–780. doi:10.2307/2445854. JSTOR 2445854. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Friedman, W. E. (1995-04-25). "Organismal duplication, inclusive fitness theory, and altruism: understanding the evolution of endosperm and the angiosperm reproductive syndrome". Proceedings of the National Academy of Sciences (باللغة الإنجليزية). 92 (9): 3913–3917. doi:10.1073/pnas.92.9.3913. ISSN 0027-8424. PMC 42072. PMID 11607532. مؤرشف من الأصل في 20 يونيو 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Friedman, William E. (1994). "The Evolution of Embryogeny in Seed Plants and the Developmental Origin and Early History of Endosperm". American Journal of Botany. 81 (11): 1468–1486. doi:10.2307/2445320. JSTOR 2445320. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Bowe, L. Michelle; Coat, Gwénaële; dePamphilis, Claude W. (2000-04-11). "Phylogeny of seed plants based on all three genomic compartments: Extant gymnosperms are monophyletic and Gnetales' closest relatives are conifers". Proceedings of the National Academy of Sciences (باللغة الإنجليزية). 97 (8): 4092–4097. doi:10.1073/pnas.97.8.4092. ISSN 0027-8424. PMC 18159. PMID 10760278. مؤرشف من الأصل في 20 سبتمبر 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Winter, Kai-Uwe; Becker, Annette; Münster, Thomas; Kim, Jan T.; Saedler, Heinz; Theissen, Günter (1999-06-22). "MADS-box genes reveal that gnetophytes are more closely related to conifers than to flowering plants". Proceedings of the National Academy of Sciences (باللغة الإنجليزية). 96 (13): 7342–7347. doi:10.1073/pnas.96.13.7342. ISSN 0027-8424. PMID 10377416. مؤرشف من الأصل في 20 يونيو 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Magallon, S.; Sanderson, M. J. (2002-12-01). "Relationships among seed plants inferred from highly conserved genes: sorting conflicting phylogenetic signals among ancient lineages". American Journal of Botany (باللغة الإنجليزية). 89 (12): 1991–2006. doi:10.3732/ajb.89.12.1991. ISSN 1537-2197. مؤرشف من الأصل في 09 ديسمبر 2019. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Chaw, Shu-Miaw; Parkinson, Christopher L.; Cheng, Yuchang; Vincent, Thomas M.; Palmer, Jeffrey D. (2000-04-11). "Seed plant phylogeny inferred from all three plant genomes: Monophyly of extant gymnosperms and origin of Gnetales from conifers". Proceedings of the National Academy of Sciences (باللغة الإنجليزية). 97 (8): 4086–4091. doi:10.1073/pnas.97.8.4086. ISSN 0027-8424. PMID 10760277. مؤرشف من الأصل في 30 نوفمبر 2018. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Zenkteler, M. (1990). "In vitro fertilization and wide hybridization in higher plants". Crit Rev Plant Sci. 9 (3): 267–279. doi:10.1080/07352689009382290. الوسيط
|CitationClass=تم تجاهله (مساعدة) -
Raghavan, V. (2005). Double fertilization: embryo and endosperm development in flowering plants (الطبعة illustrated). Birkhäuser. صفحات 17–19. ISBN 3-540-27791-9. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Campbell N.A; Reece J.B (2005). Biology (الطبعة 7). San Francisco, CA: Pearson Education, Inc. صفحات 774–777. ISBN 0-8053-7171-0. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة علم النبات
بوابة علم النبات بوابة علوم
بوابة علوم