نماذج الغشاء
قبل ظهور المجهر الإلكتروني في 1950s، والعلماء لا يعرفون بنية غشاء الخلية أو ماهي مكوناته . علماء الأحياء وغيرهم من الباحثين استخدموا أدلة غير مباشرة لتحديد صورة الغشاء قبل أن يتمكنوا فعلا من رؤيته. على وجه التحديد، كان ذلك من خلال نماذج من إرنست أوفرتون إرفينغ لانغموير، رتيير وGrendel، وDavson ودانيلي، الذي استنتج أن الأغشية تحتوي على الدهون، بروتينات، وليبيد ثنائي الطبقة. ظهور المجهر الإلكتروني، ونتائج ج. ديفيد روبرتسون، واقتراح المغني ونيكلسون، والعمل الإضافي من أنوين وهندرسون كل ذلك ساهم في تطوير نموذج الغشاء الحديث. ومع ذلك، فهم نماذج الغشاء في الماضي يلقي الضوء على تصور خصائص الغشاء في الوقت الحاضر . وبعد البحث التجريبي المكثف، سهلت نماذج الغشاء من القرن الماضي الطريق إلى نموذج الفسيفساء السائل التي يتم قبوله اليوم.
نظرية Gorter وGrendel للغشاء (1925)
Evert Gorter وFrançois Grendel (فسيولوجي هولندي) اقتربوا من اكتشاف نموذجنا الحالي لغشاءالبلازما باعتباره هيكل دهن ثنائي الطبقة. وهذه النظرية افترضت ببساطة أنه إذا كان غشاء البلازما هو طبقة ثنائية ، اذن قياس مساحة سطح طبقة أحادية من الدهون سيكون ضعف المساحة السطحية لغشاء البلازما. لفحص فرضيتهم، قاموا بتنفيذ تجربة، حيث أنهم قاموا باستخراج الدهون من عدد معروف من خلايا الدم الحمراء ( كريات الدم الحمراء ) من مصادر ثدييات مختلفة، مثل البشر، الماعز والأغنام، وغيرها، وبعد ذلك تم نشر الدهون باعتبارها طبقة أحادية في حوض Langmuir - Blodgett. وقاما بقياس المساحة الكلية لغشاء البلازما لخلايا الدم الحمراء، واستخدما طريقة Langmuir's، وقاموا بقياس مساحة طبقة أحادية من الدهون. وبمقارنة الاثنان، قاموا بحساب نسبة تقدر ب 2:1 Mono-layer of lipids:Plasma membrane. هذا يؤيد فرضيتهم، والتي أدت إلى استنتاج مفاده أن أغشية الخلايا تتكون من اثنين من الطبقات الجزيئية.[1] اقترح العالمان هيكلا لهذه الطبقة الثنائية، رؤوس قطبية محبة للماء موجهة للخارج باتجاه البيئة المائية وذيول كارهة للماء موجهة للداخل بعيدة عن المحيط المائي على جانبي الغشاء. على الرغم من أنهم وصلوا إلى الاستنتاجات الصحيحة، بعض البيانات التجريبية كانت غير صحيحة مثل سوء تقدير المساحة والضغط للدهون أحادية الطبقة وعدم اكتمال استخراج الدهون. كماأنهم فشلوا في وصف وظيفة الغشاء، وكان لديهم ايضاً افتراضات خاطئة كافتراضهم أن أغشية البلازما تتكون في معظمها من الدهون. ومع ذلك، على وجه العموم، أصبح هذا التصور لهيكل الدهن ثنائي الطبقة الافتراض الأساسي لكل تطور في الفهم الحديث لوظيفة الغشاء.[2]
نموذج Danielli وDavson بمساعدة Robertson (مابين 1940-1960)
 مقالة مفصلة: نموذج دافسون ودانيلي
مقالة مفصلة: نموذج دافسون ودانيلي
بناء على اقتراح Gorter وGrendel، نشأت شكوك حتمية على صحة وجود مجرد طبقة ثنائية بسيطة من الدهون على انها غشاء .على سبيل المثال نموذجهم عجز عن تقديم اجابات على اسئلة عن التوتر السطحي، والنفاذية، والمقاومة الكهربائية للأغشية.لذلك فإن كل من الفيزيائيHugh Davson والأحيائي James Danielli اقترحا أنه في الواقع الأغشية تحتوي بروتينات، ووفقاً لذلك فإن وجود هذه"بروتينات الغشاء"توضح ما لا يمكن الإجابة عنه بواسطة نموذج Gorter-Grendel.

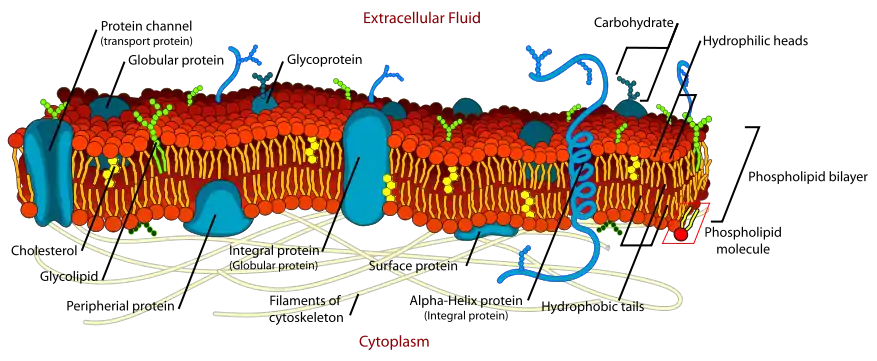
في عام 1935، اقترح Davson وDanielli أن الأغشية البيولوجية التي تتكون من دهون ثنائية الطبقة مصقولة على كلا الجانبين بصفائح رقيقة من البروتين، وقاموا بتبسيط نموذجهم إلى نظرية "pauci-molecular".[3] أوضحت هذه النظرية أن جميع الأغشية البيولوجية لديها مركز "دهني" محاط بدهون أحادية الطبقة والتي تغطيها بروتينات أحادية الطبقة. باختصار، تجلى نموذجهم على شكل "ساندويتش" بالتتابع من البروتينات والدهون والبروتينات. ألقى نموذج Davson-Danielli الضوء من جديد على فهم أغشية الخلايا، من خلال التأكيد على الدور الهام الذي تلعبه البروتينات في الأغشية البيولوجية.في 1950s، علماء الخلية تحققوا من وجود أغشية البلازما باستخدام المجهر الإلكتروني (التي يعمل بدقة عالية).واستخدم J. ديفيد روبرتسون هذا الطريقة لاقتراح نموذج الغشاء الموحّد .[4] في الأساس، اقترح أن جميع الأغشية الخلوية تشترك في بنية أساسية مماثلة،غشاء واحد. باستخدام تلطيخ المعادن الثقيلة، كما يبدو أن اقتراح روبرتسون يتوافق مع نموذج Davson-Danielli.وفقا للنمط ثلاثي الطبقات للغشاء الخلوي الذي تمت دراسته من قبل روبرتسون، اقترح أن الأغشية تتكون من طبقة ثنائية الدهون تمت تغطيتها على كلا الجانبين بصفائح رقيقة من البروتينات. وشكل هذا الاقتراح دعم كبير لاقتراح Davson وDanielli.[5]قالب:غشاء خلوي
لكن حتى مع اثبات Robertson، كان لنموذج دافسون ودانيلي تعقيدات مهمة، أهمها ان البروتينات التي تمت دراستها كانت كروية بالأساس، وبالتالي لا يمكن ان تصلح في نموذج صفائح البروتين الرقيقة.هذه الصعوبات بهذا النموذج حفزت بحوث جديدة في نظام الغشاء كما انها مهدت الطريق لنموذج الفسيفساء السائل، الذي اقترح عام 1972.
نموذج Singer وNicolson نموذج الفسيفساء السائل(1972)
- مقالة مفصلة: نموذج فسيفسائي مائع
في عام 1972، سايمور جوناثان سينجر وGarth Nicolson طوروا أفكار جديدة لبنية الغشاء.كان اقتراحهم نموذج الفسيفساء السائل ، والذي هو النموذج السائد الآن، يمتلك اثنين من الميزات الرئيسية-فسيفساء من البروتينات تترسخ في الغشاء، وطبقة ثنائية مائعة من الدهون .اقتراح الطبقة الثنائية من الدهون يتفق مع النماذج السابقة على عكس البروتينات الكروية التي تترسخ في الطبقة بدلا من الصفائح الرقيقة على السطح. وفقا للنموذج، فإن بروتينات الغشاء تنقسم إلى ثلاث فئات على أساس كيفية ارتباطها بالدهون ثنائية الطبقة: # بروتين غشائي مدمج: منغمس في الطبقة الثنائية ويرتبط في مكانه بواسطة ألفة وانجذاب الاجزاء الكارهة للماء في البروتين ناحية الذيول الكارهة للماء في الدهون الفوسفاتية في المنطقة الداخلية للطبقة. # بروتين غشائي محيطي : أكثر ألفة للماء محبة للماء، وبالتالي فهي غير - تساهمية الربط للرؤوس القطبية من الدهون الفوسفاتية وغيرها من الأجزاء المحبة للماء من بروتينات الغشاء على سطح الغشاء. # بروتين شحمي: وهو في الأساس مُحب للماء، لذلك يقع أيضا على سطح الغشاء، ويرتبط تساهمياً بجزيئات الدهون المنغرسة في الطبقة. أما بالنسبة لطبيعة الميوعة في الغشاء، فإن مكونات الدهون قادرة على التحرك بشكل موازي لسطح الغشاء وهي في حركة مستمرة، والعديد من البروتينات هي أيضا قادرة على الحركة داخل الغشاء، ومع ذلك يتم تقييد قدرتها على الحركة بسبب ارتكازها على عناصر هيكلية مثل الهيكل الخلوي على جانبي الغشاء.
بشكل عام، فإن هذا النموذج يوضح معظم الانتقادات ل نموذج Davson–Danielli، حيث أنه يلغي الحاجة لاستيعاب بروتينات الغشاء في الطبقات السطحية الرقيقة، كما أنه يوضح التغير في معدل البروتين / الدهون في الأغشية المختلفة، مما يعني ببساطة أن الأغشية المختلفة تختلف في كمية البروتين التي تحتويها، كما أنه يوضح تلاؤم مجموعة الرؤوس الدهنية على سطح الغشاء مع حساسيتها لهضم الفسفوليباز .أيضا، ميوعة الدهون ثنائية الطبقة وامتزاج مكوناتها داخل الغشاء يجعل من السهل رؤية تنقل وحركة كل من الدهون والبروتينات.
نظرية Henderson وUnwin للغشاء
درس كل من Henderson وUnwin الغشاء الأرجواني بواسطة المجهر الإلكتروني، وذلك باستخدام طريقة لتحديد الأشكال المتوقعة للعينات البلورية الغير مصبوغة. من خلال تطبيق طريقة انحراف العينات، وباستخدام المبادئ التي طرحها كل من DeRosier وKlug لمجموعة من الآراء ووجهات النظر ثنائية الأبعاد، فحصلوا على خريطة ثلاثية الأبعاد للغشاء بنسبة 7 ؛ وثبات Å.الخريطة تكشف عن مكان وجود مكونات البروتين والدهون، وترتيب سلاسل الببتيد داخل كل جزيء بروتين، والعلاقة بين جزيئات البروتين في الشبكة الشعرية.[6]
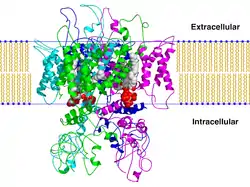
الصور المجهرية عالية الدقة للصفائف البلورية لبروتينات الغشاء، التي اتخذت بجرعة منخفضة من الإلكترونات للحد من أضرار الإشعاع، تم استغلالها لتحديد هيكل ثلاثي الأبعاد بواسطة تحويل فورييه.الدراسات التي أجريت مؤخرا على الفئران الملونة الكبدية والتي اعتمدت على أن تخضع تقاطعات الفجوة لإعادة بناء فورييه ثلاثي الأبعاد (بجرعة منخفضة من صورة مجهريةs) تشير إلى أن يتم ترتيب الوحدات الفرعية الستة للبروتين في أسطوانة مائلة قليلا بشكل عرضي، داخل قناة واسعة 2 nm في المنطقة الخارج خلوية .وكانت أبعاد القناة داخل الغشاء ضيقة، لكن لا يمكن توسيعها(Unwin and Zampighi, 1980), عدا عن أن حركة راديكالية صغيرة من الوحدات الفرعية في النهايات السيتوبلازمية يمكن أن تقلل من الانحراف المماسي للوحدة الفرعية إلى المحور ستة أضعاف وإغلاق القناة...[7]
مزيد من التفاصيل في التنظيم الجزيئي يجب أن تظهر كأكثر أساليب الإعدادالمتاحة، بحيث يتم الحصول على صور عالية الدقة ثلاثية الأبعاد مقارنة بالأغشية الأرجوانية التي تم الحصول عليها، باستخدام إجراءات مبتكرة لتحليل صورة المصفوفات المتكررة البيولوجية الجزيئات، عند دمج البيانات من جرعة منخفضة للصور الإلكترونية وعينات الانكسار المجمعة، Henderson وUnwin عام (1975), جددوا صورة ثلاثية الأبعاد للغشاء الأرجواني بقياس 0.7 nm.استخدم تضمين الغلوكوز لتخفيف ضرر الجفاف والجرعات المنخفضة(< 0.5 e/A*) للحد من أضرار الإشعاع، وتم حفظ الصور المجهرية الالكترونية للاغشية الغير مصبوغة وسُجلت على انها المصدر الوحيد للمقارنة واتضح ان حالة المقارنة هذه ضعيفة بسبب التعمق في التركيز .
وفي تجربتهم، وجد كل من Unwin وHenderson أن البروتين يمتد إلى جانبي الدهون ثنائية الطبقة ويتكون من سبعة لفائف من نوع ألفا مجمعة في رزمة طولها يتراوح ما بين 1–1.2 nm؛ إلى 3.5–4.0 nm ، ويسري بشكل عمودي على سطح الغشاء، حيث يتم تنظيم الجزيئات حول محور 3 أضعاف ومساحة بعرض 2 nm؛ مركزها مليء بالدهون. ويمثل هذا العمل الرائع أهم خطوة إلى الأمام حتى الآن، كما وفر لنا في المرة الأولى زودنا بنية البروتين المكمل للغشاء . توفر سلسلة من الأحماض الأمينية، مع معلومات حول كثافة انتشار الإلكترون من عمل Henderson وUnwin، قد حفز جهود جديدة لبناء نموذج (Engleman et al., 1980) ليتناسب مع معلومات تسلسل جرثومية رودوبسين داخل سلسلة من شرائح-α الحلزونية.
انظر أيضاً
- نموذج دانيل ودافسون
- نموذج فسيفسائي مائع
- بيولوجيا الخلية
- نظرية الخلية
- نظرية-إم
- بروتين غشائي
مراجع
- "Membrane – An Introduction" (PDF). Wiley-VCH. مؤرشف من الأصل (PDF) في 8 مايو 2016. اطلع عليه بتاريخ 09 أكتوبر 2015. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Becker's World of the Cell (الطبعة 8th). University of Wisconsin-Madison: Jeff Hardin. 2012. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Robertson, J. David. "Membrane Structure" (PDF). jcb.rupress.org. jcb.rupress.org. مؤرشف من الأصل (PDF) في 27 أبريل 2020. اطلع عليه بتاريخ 09 أكتوبر 2015. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Heuser, John E. "In Memory of J.David Robertson" (PDF). heuserlab.wustl.edu. heuserlab.wustl.edu. مؤرشف من الأصل (PDF) في 8 أكتوبر 2018. اطلع عليه بتاريخ 08 أكتوبر 2015. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Hardin, Jeff; Kleinsmith, Lewis J.; Bertoni, Gregory; Becker, Wayne M. (2012). World of the Cell (الطبعة Eighth). US: Pearson Benjamin Cummings. صفحات 158–163. اطلع عليه بتاريخ 09 أكتوبر 2015. الوسيط
|CitationClass=تم تجاهله (مساعدة)صيانة CS1: التاريخ والسنة (link) - R. Henderson & P. N. T. Unwin (September 4, 1975). "Three-dimensional model of purple membrane obtained by electron microscopy". Nature. Cambridge: MRC Laboratory of Molecular Biology. 257: 28, 29. doi:10.1038/257028a0. مؤرشف من الأصل في 27 أبريل 2020. الوسيط
|CitationClass=تم تجاهله (مساعدة) - Malhotra, S. K. (1983). The Plasma membrane. Canada: John Wiley & Sons. صفحات 3, 92, 9 95. الوسيط
|CitationClass=تم تجاهله (مساعدة)
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي