بروتين تاو
توجد ستة أشكال من بروتين تاو في أنسجة الدماغ البشري، ويتم التمييز بينها من خلال عدد نطاقات البرويتين .ثلاثة من اشكال البروتين لديها ثلاثة نطاقات للربط والثلاثة الأخرى لديها أربعة نطاقات للربط . ويكون مكان انطقة الربط هذه في النهاية الكربوكسيلية من البروتين وتكون مشحونة بشحنة موجبة ( ذلك ليتم الارتباط مع الانيببات الدقيقة سالبة الشحنة ) . اشكال التاو التي تحوي على اربعة انطقة للارتباط تكون احسن من تلك التي تحوي فقط على ثلاثة انطقة للربط وذلك لانه تزيد من استقرار الانيببات الدقيقة . وهذه الاشكال المتعددة من برتين تاو نتجت بسبب الوصل المتبادل للإكسون 2 , 3 و 10 في جين تاو .
| Microtubule associated protein tau | |||||||
|---|---|---|---|---|---|---|---|
| المعرفات | |||||||
| الأسماء المستعارة | PHF-tau, microtubule-associated protein tau, neurofibrillary tangle protein, tau proteins, protein phosphatase 1, regulatory subunit 103, paired helical filament-tau, G protein beta1/gamma2 subunit-interacting factor 1, MAPT, tau protein | ||||||
| معرفات خارجية | |||||||
| نمط التعبير عن الحمض النووي الريبوزي | |||||||
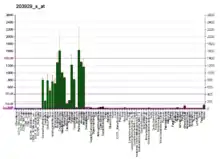 | |||||||
| المزيد من بيانات التعبير المرجعية | |||||||
| أورثولوج | |||||||
| الأنواع | الإنسان | الفأر | |||||
| أنتريه | n/a | ||||||
| Ensembl | n/a | n/a | |||||
| يونيبروت |
|
| |||||
| RefSeq (مرسال ر.ن.ا.) |
|
| |||||
| RefSeq (بروتين) |
|
| |||||
| الموقع (UCSC | n/a | ||||||
| بحث ببمد | n/a | ||||||
| ويكي بيانات | |||||||
| |||||||
الشكل :
تاو هو فوسفوبروتين مع 79 سيرين المحتملة (Ser) وثريونين (Thr) مواقع الفسفرة على أطول شكل من اشكال تاو. تم الإبلاغ عن الفسفرة في حوالي 30 من هذه المواقع في بروتينات تاو العادية .[1]
وينظم فسفرة البروتين تاو من قبل مجموعة من كيناز، بما في ذلك ( PKN ) ، وهو سيرين / ثريونين كيناز. عندما يتم تنشيط( PKN ) ، يتم فسفرة بروتين تاو، مما يؤدي إلى تفكيك الانيببات الدقيقة .[2]
ويتم تنظيم فسفرة البروتين تاو مع تطور المراحل الجنينية . على سبيل المثال، يكون البروتين تاو في المراحل الجنينية أكثر فسفوريلية في الجهاز العصبي المركزي من البروتين تاو عند الكبار.[3] درجة الفسفرة في جميع اشكال البروتين تاو الستة تتناقص مع التقدم في السن بسبب تفعيل الفوسفاتاسيس.[4] مثل كيناز، فوسفاتاسيس أيضا تلعب دورا في تنظيم الفسفرة من تاو. على سبيل المثال، PP2A و PP2B على حد سواء موجودة في أنسجة الدماغ ولديها القدرة على ديفوسفوريلات Ser396 .[5] ربط هذه الفوسفاتاتيس إلى تاو يؤثر على الترابط ما بين تاو والانيببات الدقيقة .
بروتينات تاو (أو بروتينات τ، وهو حرف يوناني) هي البروتينات التي تساعد على استقرار الأنابيب الدقيقة. فهي وفيرة في الخلايا العصبية في الجهاز العصبي المركزي وأقل شيوعا في أماكن أخرى، كما يتم التعبير عنها أيضا في مستويات منخفضة جدا في الجهاز العصبي المركزي في الخلايا النجمية وخلايا الأوليغوديندرايت .[6]
ترتبط الأمراض والخرف في الجهاز العصبي مثل مرض الزهايمر ومرض باركنسون [7] ببروتينات تاو التي أصبحت مختلة ولم تعد قادرة على توفير الاستقرار للانابيب الدقيقة بالشكل صحيح.
بروتينات تاو هي نتاج الربط البديلة من جين واحد في الانسان هو( MAPT) ( microtubule -associated protein tau) ويقع على الكروموسوم 17.[8][9]
تم تحديد بروتينات تاو في عام 1975 باعتبارها بروتينات مستقرة للحرارة ضرورية لتجميع الأنابيب الدقيقة [10][11]، وقد تبين أن لها خصائص البروتينات .[12]
الوظيفة
البروتين تاو هو من البروتينات عالية الذائبية المرتبطة بالانيببات الدقيقة (MAP) . في الانسان، توجد هذه البروتينات في الغالب في الخلايا العصبية مقارنة بالخلايا غير العصبية. واحدة من المهام الرئيسية لبروتين تاو هي المحافظة على استقرار الأنابيب الدقيقة في المحور العصبي . يوجد في الجهاز العصبي بروتينات (MAPs ) اخرى وتؤدي نفس الوظيفة، كما اقترح من قبل تجربة الفئران ( tau knockout mice ) التي لم تظهر تشوهات اي في نمو الدماغ - ربما بسبب التعويض في نقص تاو من قبل( MAPs ) أخرى .[13] البروتين تاو غير موجود في التشعبات وينشط في الأجزاء البعيدة من المحاور حيث أنه يوفر استقرار الأنيببات والمرونة أيضا حسب الحاجة. هذا يتناقض مع البروتينات MAP6 (STOP في الأجزاء القريبة من المحاور، والتي في دورها تؤمن الأنابيب الدقيقة و MAP2 التي تعمل على استقرار الأنابيب الدقيقة في التشعبات.
تتفاعل بروتينات تاو مع توبولين لتحقيق الاستقرار في الأنابيب الدقيقة وتعزيز تجميع الأنبوبة في الأنابيب الدقيقة.[11] بروتين تاو لديه طريقتان للسيطرة على استقرار أنيبيب: إيسوفورمز والفسفرة .
مراجع
- Billingsley ML, Kincaid RL (May 1997). "Regulated phosphorylation and dephosphorylation of tau protein: effects on microtubule interaction, intracellular trafficking and neurodegeneration". Biochem. J. 323 (3): 577–91. ببمد سنترال 1218358. ببمد 9169588. نسخة محفوظة 30 سبتمبر 2002 على موقع واي باك مشين.
- Taniguchi T, Kawamata T, Mukai H, Hasegawa H, Isagawa T, Yasuda M, Hashimoto T, Terashima A, Nakai M, Mori H, Ono Y, Tanaka C (March 2001). "Phosphorylation of tau is regulated by PKN". J. Biol. Chem. 276 (13): 10025–31. معرف الوثيقة الرقمي:. ببمد 11104762. نسخة محفوظة 04 مايو 2017 على موقع واي باك مشين.
- Kanemaru K, Takio K, Miura R, Titani K, Ihara Y (May 1992). "Fetal-type phosphorylation of the tau in paired helical filaments". J. Neurochem. 58 (5): 1667–75. معرف الوثيقة الرقمي:. ببمد 1560225. نسخة محفوظة 04 مايو 2017 على موقع واي باك مشين.
- Mawal-Dewan M, Henley J, Van de Voorde A, Trojanowski JQ, Lee VM (December 1994). "The phosphorylation state of tau in the developing rat brain is regulated by phosphoprotein phosphatases". J. Biol. Chem. 269 (49): 30981–7. ببمد 7983034. نسخة محفوظة 04 مايو 2017 على موقع واي باك مشين.
- Matsuo ES, Shin RW, Billingsley ML, Van deVoorde A, O'Connor M, Trojanowski JQ, Lee VM (October 1994). "Biopsy-derived adult human brain tau is phosphorylated at many of the same sites as Alzheimer's disease paired helical filament tau". Neuron. 13 (4): 989–1002. معرف الوثيقة الرقمي:. ببمد 7946342. نسخة محفوظة 04 مايو 2017 على موقع واي باك مشين.
- Shin RW, Iwaki T, Kitamoto T, Tateishi J (May 1991). "Hydrated autoclave pretreatment enhances tau immunoreactivity in formalin-fixed normal and Alzheimer's disease brain tissues". Lab. Invest. 64 (5): 693–702. ببمد 1903170. نسخة محفوظة 02 يوليو 2017 على موقع واي باك مشين.
- Lei P, Ayton S, Finkelstein DI, Adlard PA, Masters CL, Bush AI (November 2010). "Tau protein: relevance to Parkinson's disease". Int J Biochem Cell Biol. 42 (11): 1775–1778. معرف الوثيقة الرقمي:. ببمد 20678581. نسخة محفوظة 29 مارس 2015 على موقع واي باك مشين.
- Goedert M, Wischik CM, Crowther RA, Walker JE, Klug A (June 1988). "Cloning and sequencing of the cDNA encoding a core protein of the paired helical filament of Alzheimer disease: identification as the microtubule-associated protein tau". Proc. Natl. Acad. Sci. U.S.A. 85 (11): 4051–5. معرف الوثيقة الرقمي:. ببمد سنترال 280359. ببمد 3131773. نسخة محفوظة 15 يوليو 2017 على موقع واي باك مشين.
- Goedert M, Spillantini MG, Jakes R, Rutherford D, Crowther RA (October 1989). "Multiple isoforms of human microtubule-associated protein tau: sequences and localization in neurofibrillary tangles of Alzheimer's disease". Neuron. 3 (4): 519–26. معرف الوثيقة الرقمي:. ببمد 2484340. نسخة محفوظة 18 أكتوبر 2017 على موقع واي باك مشين.
- Weingarten MD, Lockwood AH, Hwo SY, Kirschner MW (May 1975). "A protein factor essential for microtubule assembly". Proc. Natl. Acad. Sci. U.S.A. 72 (5): 1858–62. معرف الوثيقة الرقمي:. ببمد سنترال 432646. ببمد 1057175. نسخة محفوظة 07 مارس 2016 على موقع واي باك مشين.
- Cleveland DW, Hwo SY, Kirschner MW (October 1977). "Purification of tau, a microtubule-associated protein that induces assembly of microtubules from purified tubulin". Journal of Molecular Biology. 116 (2): 207–225. معرف الوثيقة الرقمي:. ببمد 599557. نسخة محفوظة 24 أبريل 2017 على موقع واي باك مشين.
- Cleveland DW, Hwo SY, Kirschner MW (October 1977). "Physical and chemical properties of purified tau factor and the role of tau in microtubule assembly". Journal of Molecular Biology. 116 (2): 227–247. معرف الوثيقة الرقمي:. ببمد 146092. نسخة محفوظة 25 فبراير 2016 على موقع واي باك مشين.
- Harada A, Oguchi K, Okabe S, Kuno J, Terada S, Ohshima T, Sato-Yoshitake R, Takei Y, Noda T, Hirokawa N (June 1994). "Altered microtubule organization in small-calibre axons of mice lacking tau protein". Nature. 369 (6480): 488–91. معرف الوثيقة الرقمي:. ببمد 8202139. نسخة محفوظة 24 أبريل 2017 على موقع واي باك مشين.
روابط خارجية
- tau Proteins في المَكتبة الوَطنية الأمريكية للطب نظام فهرسة المواضيع الطبية (MeSH).
- GeneReviews/NCBI/NIH/UW entry on MAPT-Related Disorders
- MR scans of variant CJD CSF Tau positive man
 بوابة طب
بوابة طب بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية
10.1074/jbc.M007427200 10.1111/j.1471-4159.1992.tb10039.x 10.1016/0896-6273(94)90264-X 10.1016/j.biocel.2010.07.016 10.1073/pnas.85.11.4051 10.1016/0896-6273(89)90210-9 10.1073/pnas.72.5.1858 10.1016/0022-2836(77)90213-3 10.1016/0022-2836(77)90214-5 10.1038/369488a0